Бм список ниш: 500 ниш от Бизнес
Как выбрать нишу в инфобизнесе
Алгоритм того, как выбрать нишу в инфобизнесе можно рассмотреть, применив практику «Бизнес-Молодость». Хотя Петр Осипов и компания не учат тому, как создавать именно инфобизнес, но их наработки по запуску своего дела можно с успехом адаптировать применительно и к информационным интернет-проектам.
Азбука инфобизнеса: выбор ниши
Для того чтобы разобраться в том, как выбрать нишу в инфобизнесе, мы будем использовать вот такую таблицу:
Таблица №1. Ниша инфобизнеса: базовая таблица при выборе
Начнем наш анализ с самого идеального варианта. На примере таблицы №2 рассмотрим, как именно работает алгоритм выбора ниши в инфобизнесе.
Таблица №2. Лучшие ниши инфобизнеса
Самые лучшие ниши инфобизнеса (алгоритм выбора отмечен зеленым) — это те, работая с которыми можно рассчитывать на одновременное соблюдение трех условий: Вы — эксперт в выбранной нише; она Вам нравится и на нее есть спрос среди посетителей Интернета.
При других обстоятельствах у человека, который размышляет о том, как создать инфобизнес, выбор может быть связан со следующими вариантами.
- Новичок инфобизнеса не считает себя экспертом ни в какой области. В этой ситуации смотрим правую часть таблицы №1, выделенную оранжевым. Путь в инфобизнес для него не закрыт, но перед запуском проекта появляется промежуточный этап, связанный с обретением статуса эксперта в выбранной нише или с продюсированием инфобизнеса (подробнее об этом поговорим в других статьях блога «Законы ИнфоБизнеса»).
- Новичок инфобизнеса является специалистом в деле, которым ему не нравится заниматься. В этом случае все равно нужно выяснить, есть ли в этой нише инфобизнеса спрос. Если да, ее следует внести в список для тестирования. После этого нужно действовать по алгоритму, описанному в пункте №1.


- Новичок инфобизнеса не является экспертом, а та ниша, которую он хотел бы освоить, не востребована в сети Интернет. В таком случае следует тестировать разные ниши и, выбрав те, на которые есть спрос, запускать проекты, используя продюсирование инфобизнеса.
Таблица №3. Прибыльные ниши инфобизнеса
Недостатком такого варианта является то, что в процессе развития инфобизнеса его владельцу придется в большей мере мотивировать себя в тех областях, которые ему, возможно, не нравятся. Именно поэтому такие ниши можно назвать прибыльными, но ни лучшими.
- Новичок инфобизнеса — эксперт в деле, которое в сети Интернет не востребовано и он не готов осуществлять продюсирование инфобизнеса. Такому человеку следует отказаться от намерения создать инфобизнес. Если же он будет продолжать упорно строить онлайн-проект, то столкнется с ситуацией, представленной ниже:
Таблица №4. Ниша инфобизнеса, в которой нет смысла работать
Как выбрать нишу в инфобизнесе: выводы
Итак, чтобы выбрать нишу в инфобизнесе следует принимать во внимание свой уровень экспертности и желание работать, востребованность ниши среди пользователей Интернета и готовность осуществлять продюсирование инфобизнеса. Также не нужно закрывать глаза на те ситуации, когда онлайн-проект развивать нет смысла. Применение этих советов поможет успешно начать инфобизнес с нуля.
Подписывайтесь на обновление блога «Законы ИнфоБизнеса» чтобы своевременно получать информацию обо всех аспектах, связанных с правильным выбором ниши в инфобизнесе.
Как стартовать в бизнесе с нуля без денег? 15 идей и список конкретных шагов | Бизнес на все 💯
В данном выпуске организаторы Бизнес Молодости помогают переместиться от раздумий к ситуации, когда во вновь созданной компании появляются клиенты.
Наличие желающих приобрести товар или услугу сопровождается вашей возможностью исполнить по отношению к ним обязательства, которые впоследствии превратятся в деньги.
Именно этот процесс называется бизнесом – он рождается с первой сделки.
Идеи для старта – составьте длинный список ниш – для этого процесса используйте базовые или приобретенные навыки:
- Образование – дает преимущество перед другими людьми, область знаний направит на написание 5-6 ниш.
- Семья – это поле для взаимосвязи деятельности членов семьи с вашей деятельностью.
- Друзья и ближайшее окружение – поле похожее на семью для развития совместного бизнеса.
- Естественный интерес – хобби, естественное направление внимание без особых усилий, «чем больше времени вы вкладываете в занятие, тем выше ваша экспертность».
- Жизненный опыт – любой жизненный опыт послужит развитию идеи для ниши.
- Навыки – способность сделать полезные вещи в любой момент времени.
- Источник вдохновения – перенятые бизнес идеи с другой страны.
- Ответ на боль – рынок не в состоянии удовлетворить текущую потребность человека.
- Невидимые ниши – редко востребованные ниши, которыми пользуются определенное количество потребителей в узком кругу.
- Существующий спрос – проверка существующего спроса на Worstat.Yandex.ru, который помогает настроить поиск сайта компании по ключевым словам.
- Китай – посещение выставок или площадок китайских товаров способствует выбору нестандартных ниш.
- Отраслевые выставки – поставщики и производители на подобных мероприятиях изыскивают продавцов.
- Приобретение новых навыков – обучение новым профессиям позволяет уверенней чувствовать себя на рынке труда.
- Взгляды и убеждения – это направление всегда способствует развитию хобби и как следствие выбору очередной ниши.
- Очки Бизнес Молодость – в подобных очках все окружающие предметы станут возможными нишами.


Смотрите полностью все идеи на видео, источник — БМ:
Текущие закупки
Тип закупкиОтборКонкурсАукционЗапрос предложенийЗапрос котировокМаркетинговые исследования
Категория — выбрать все 1 — — снять выделение -Прочие строительно-монтажные работы — выбрать все 3 — — снять выделение -Поставка лицензий и ПОИнформационные технологииТехническая поддержка оборудования и ПО — выбрать все 7 — — снять выделение -Запчасти и комплектующие к технике
(Поставка МТР: производственных)Спецодежда, спецобувь и другие сизМТРЗапчасти к оборудованию механическому
Канцтовары, офисная техн.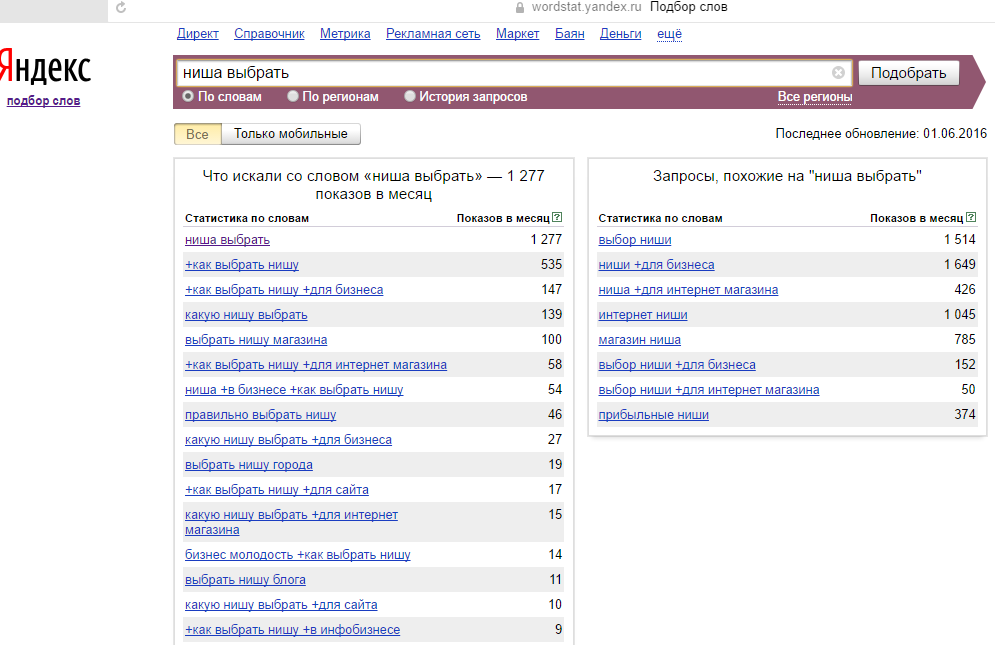
Место оказания работ/услугМоскваСанкт-ПетербургНовосибирскЕкатеринбургНижний НовгородКазаньСамараОмскЧелябинскРостов-на-ДонуУфаВолгоградПермьКрасноярскВоронежСаратовКраснодарТольяттиИжевскУльяновскБарнаулВладивостокЯрославльИркутскТюменьМахачкалаХабаровскБалейкинское м/р (ООО «Газпромнефть-Оренбург»)ОренбургНовокузнецкКемеровоХанты-МансийскНовороссийскКрайнее м/р (Филиал «Газпромнефть-Муравленко» ОАО «Газпромнефть-ННГ»)Cеверо-восточная часть Пальяновской площади (ООО «Газпромнефть-Хантос»)Арчинское м/р (ООО «Газпромнефть-Восток»)Аяшский лицензионный участокВалынтойское (Филиал «Газпромнефть-Муравленко» ОАО «Газпромнефть-ННГ»)Воргенское нефтяное м/р (АО «Газпромнефть-ННГ»)Восточно-Мессояхское и Западно-Мессояхское м/р (АО «Мессояханефтегаз»)Восточно-Мессояхское м/р (АО «Мессояханефтегаз»)Восточный участок Оренбургского НГКМ (ООО «Газпромнефть-Оренбург»)Вынгапуровское нефтегазовое м/р (АО «Газпромнефть-ННГ»)Вынгаяхинское (Филиал «Газпромнефть-Муравленко» ОАО «Газпромнефть-ННГ»)Вынгаяхинское, Еты-Пуровское, Валынтойское (Филиал «Газпромнефть-Муравленко» ОАО «Газпромнефть-ННГ»)Долгинское месторождениеЕты-Пуровское месторождение (Филиал «Газпромнефть-Муравленко» ОАО «Газпромнефть-ННГ»)Западно-Лугинецкое м/р (ООО «Газпромнефть-Восток»)Западно-Мессояхское м/р (АО «Мессояханефтегаз»)Западно-Ноябрьское нефтяное м/р (АО «Газпромнефть-ННГ»)Зимнее м/р (ООО «Газпромнефть-Хантос»)Игнялинский и Тымпучиканский лицензионные участкиИгнялинский лицензионный участокКапитоновское м/р (ООО «Газпромнефть-Оренбург»)Карамовское нефтяное м/р (АО «Газпромнефть-ННГ»)Кемеровская нефтебаза ЗАО «Газпромнефть-Терминал»Красноленинское м/р (ООО «Газпромнефть-Хантос»)Нижнелугинецкое м/р (ООО «Газпромнефть-Восток»)Новогоднее нефтегазовое м/р (АО «Газпромнефть-ННГ»)Новопортовское месторождениеОрехово-Ермаковское м/р (ООО «Газпромнефть-Хантос»)Пограничное нефтяное м/р (АО «Газпромнефть-ННГ»)Приразломное месторождениеПякяхинское месторождениеРавнинное нефтегазовое м/р (АО «Газпромнефть-ННГ»)Спорышевское нефтяное м/р (АО «Газпромнефть-ННГ»)Средне-Итурское нефтяное м/р (АО «Газпромнефть-ННГ»)Уватский район (ООО «Газпромнефть-Хантос»)Урманское м/р (ООО «Газпромнефть-Восток»)Холмистое нефтегазовое м/р (АО «Газпромнефть-ННГ»)Холмогорское нефтяное м/р (АО «Газпромнефть-ННГ»)Царичанское м/р (ООО «Газпромнефть-Оренбург»)Чатылькинское нефтегазовое м/р (АО «Газпромнефть-ННГ»)Шингинское м/р (ООО «Газпромнефть-Восток»)Юго-Западная часть Крапивинского м/р (ООО «Газпромнефть-Восток»)Южная лицензионная территория Приобского м/р (ООО «Газпромнефть-Хантос»)Южно-Киняминское м/р (ООО «Газпромнефть-Хантос»)Южно-Приобское м/р (ООО «Газпромнефть-Хантос»)Воргенское нефтяное м/рЯрайнерское нефтегазовое м/р (АО «Газпромнефть-ННГ»)
| Аннотация |
Актуальность и цели. В условиях Самарской Луки два вида – P. nathusii и E. nilssonii – обитают совместно и демонстрируют сходные экологические потребности, что должно обусловливать определенный уровень их конкурентных отношений. Целью работы было изучение трофической экологии этих видов, а также выявление уровня перекрывания их экологических ниш в условиях Самарской Луки. Рацион питания устанавливали путем анализа фрагментов насекомых в экскрементах. Результаты и выводы. За время проведения работ нами было отловлено 1469 особей P. nathusii и 744 особей E. nilssonii. В макроместообитаниях наибольшая встречаемость P.  nathusii выявлена в островной пойме и береговой зоне, а E. nilssonii – в лиственных лесах, произрастающих по оврагам северных склонов Жигулевских гор. В микроместообитаниях, связанных с использованием определенных типов охотничьих пространств и охотничьего поведения, виды проявляют заметное сходство. Принципиальные отличия сводятся в использовании расширенного диапазона охотничьих пространств и проявляются в структуре пищевого рациона. В питании P. nathusii выявлено восемь категорий кормов, принадлежащих семи отрядам насекомых, у E. nilssonii – девять категорий, принадлежащих восьми отрядам. В диете P. nathusii преобладают Chironomidae, а у E. nilssonii – Lepidoptera. В целом для P. nathusii характерна меньшая ширина трофической ниши и более выраженная специализация в питании по сравнению со стратегией E. nilssonii. Более широкая ниша, вероятно, дает для последнего вида одно из преимуществ в конкуренции за ресурсы при доминировании по численности первого. Уровень перекрывания трофических ниш у обоих видов невелик. nathusii выявлена в островной пойме и береговой зоне, а E. nilssonii – в лиственных лесах, произрастающих по оврагам северных склонов Жигулевских гор. В микроместообитаниях, связанных с использованием определенных типов охотничьих пространств и охотничьего поведения, виды проявляют заметное сходство. Принципиальные отличия сводятся в использовании расширенного диапазона охотничьих пространств и проявляются в структуре пищевого рациона. В питании P. nathusii выявлено восемь категорий кормов, принадлежащих семи отрядам насекомых, у E. nilssonii – девять категорий, принадлежащих восьми отрядам. В диете P. nathusii преобладают Chironomidae, а у E. nilssonii – Lepidoptera. В целом для P. nathusii характерна меньшая ширина трофической ниши и более выраженная специализация в питании по сравнению со стратегией E. nilssonii. Более широкая ниша, вероятно, дает для последнего вида одно из преимуществ в конкуренции за ресурсы при доминировании по численности первого. Уровень перекрывания трофических ниш у обоих видов невелик.
|
Список литературы |
1. Уиттекер, Р. Сообщества и экосистемы / Р. Уиттекер. – М. : Прогресс, 1980. – 327 с. |
 Поволжский регион. Естественные науки
Поволжский регион. Естественные науки – М. : Мир, 1986. – Т. 2. – 375 с.
– М. : Мир, 1986. – Т. 2. – 375 с. – С. 112–237.
– С. 112–237. G. Smirnov, V. P. Vekhnik // Biology Bulletin. – 2014. – Vol. 41, № 1. – P. 60–70.
G. Smirnov, V. P. Vekhnik // Biology Bulletin. – 2014. – Vol. 41, № 1. – P. 60–70. McAney, C. Shiel, C. Sullivan, J. Fairley // The Mammal Society. – 1991. – № 14. – 48 p.
McAney, C. Shiel, C. Sullivan, J. Fairley // The Mammal Society. – 1991. – № 14. – 48 p.
[Бизнес Молодость] ЦЕХ 29 (2019)
Команда сайта FreeKurses.COM с БОЛЬШИМ УВАЖЕНИЕМ относится к каждому посетителю нашего сайта. Мы стремимся защищать Вашу конфиденциальность и уважать её. Если у Вас есть вопросы о Вашей личной информации, пожалуйста, свяжитесь с нами.
Какую информацию о Вас мы собираем
Типы данных, которые мы собираем и обрабатываем включают в себя следующее:
- Ваше реальное имя или никнейм на сайте.
- Ваш адрес электронной почты.
- Ваш IP-адрес.
Дополнительные данные могут быть собраны, если Вы решите поделиться ими, например, если Вы заполните поля в своём профиле.
Мы собираем некоторую или всю эту информацию в следующих случаях:
- Вы регистрируетесь как участник на этом сайте.
- Вы заполняете нашу форму обратной связи.
- Вы просматриваете этот сайт. Смотрите пункт «Политика в отношении файлов cookie» ниже.
- Вы заполняете поля в своём профиле.

Как используется Ваша личная информация
Мы можем использовать Вашу личную информацию в следующих целях:
- Для того, чтобы сделать Вас зарегистрированным участником нашего сайта, чтобы Вы могли добавлять и получить контент.
- Мы можем использовать Ваш адрес электронной почты, чтобы информировать Вас об активности на нашем сайте.
- Ваш IP-адрес записывается при выполнении определённых действий на нашем сайте. Он никогда не отображается публично!
Другие варианты использования Вашей личной информации
В дополнение к оповещениям Вас об активности на нашем сайте, время от времени мы можем связываться со всеми пользователями посредством рассылки на электронную почту или объявлений для того, чтобы рассказать им какую-либо важную информацию. Вы можете отказаться от подобных рассылок в своём профиле.
Вы можете отказаться от подобных рассылок в своём профиле.
В ходе Вашего использования нашего сайта мы можем собирать информацию, не связанную с личными сведениями о Вас. Эти данные могут содержать техническую информацию о браузере или типе устройства, которые Вы используете. Эта информация будет использоваться исключительно для целей аналитики и отслеживания количества посетителей нашего сайта.
Обеспечение безопасности данных
Мы стремимся обеспечить безопасность любой информации, которую Вы нам предоставляете. Чтобы предотвратить несанкционированный доступ или раскрытие информации, мы добавили подходящие меры и процедуры для защиты и обеспечения безопасности информации, которую мы собираем.
Политика в отношении файлов cookie
Cookie — это небольшие текстовые файлы, которые будут установлены нашим сайтом на Ваш компьютер, они позволят нам предоставлять определённые функции на нашем сайте, такие как возможность входа в систему или запоминание определённых предпочтений.
Более подробную информацию о cookie Вы можете прочитать на этой странице.
Права
Вы имеете право получить доступ к личным данным, которые у нас имеются о Вас, или получить их копию. Для этого свяжитесь с нами. Если Вы считаете, что информация, которую мы о Вас имеем является неполной или неточной, то Вы можете попросить нас дополнить или исправить эту информацию.
Вы также имеете право запросить удаление Ваших личных данных. Пожалуйста, свяжитесь с нами, если Вы хотите, чтобы мы удалили Ваши личные данные.
Принятие политики
Дальнейшее использование нашего сайта означает Ваше полное согласие с этой политикой. Если Вы не согласны с нашей политикой, то пожалуйста, не используйте данный сайт. При регистрации мы также запросим Ваше явное согласие с политикой конфиденциальности.
Изменения в политике
Мы можем вносить изменения в данную политику в любое время. Вам может быть предложено ознакомиться и повторно принять информацию в этой политике, если она изменится в будущем.
Прайс-лист
Строительные работы
Кладка перегородок из пазогребневых блоков или пеноблоков толщиной до 100 мм | 450 р |
Устройство металлической перемычки дверного проема | 300 р |
Кладка перегородок из пеноблоков толщиной 150 мм | 675 р |
Кладка перегородок из пеноблоков толщиной 200 мм | 900 р |
Кладка сантехнического короба из блоков | 2500 р |
Закладка проемов из пазогребневых блоков или пеноблоков толщиной до 100 мм | 700 |
Закладка перемычек дверных проемов блоками | 100 |
Выравнивание подоконной части раствором | 450 |
Кладка перегородок из кирпича (1/2 кирпича) | 900 |
Кладка перегородок из кирпича (1 кирпич) | 1800 |
Устройство экрана ванны из блоков | 1500 |
Устройство подиума из бетона до 100 мм | 3200 |
Устройство поддона душевой кабины из бетона | 4200 |
Установка стеклоблока | 200 |
Устройство структуры стеклоблоков | 1800 |
Заделка швов стеклоблоков | 200 |
Гипсокартонные работы |
|
Устройство ГКЛ потолков 1-уровневых (1 слой ГКЛ) | 1000 |
Устройство ГКЛ потолков 1-уровневых (2 слоя ГКЛ) | 1200 |
Устройство ГКЛ потолков 2-уровневых (1 слой ГКЛ) (прямые углы без ниш и скрытых подсветок) | 1200 |
Устройство ГКЛ потолков 2-уровневых (2 слоя ГКЛ) (прямые углы без ниш и скрытых подсветок) | 1500 |
Устройство ГКЛ потолков сложных фигурных 2-уровневых (1 слой ГКЛ) | 2500 |
Устройство ГКЛ потолков сложных фигурных 3-уровневых (1 слой ГКЛ) | 3000 |
Обшивка стен ГКЛ по металл. | 500 |
Обшивка стен ГКЛ по металл. каркасу (в 2 слоя) | 700 |
Установка лючка (металл, пластик) | 400 |
Установка люка-невидимки под окраску | 1000 |
Установка экрана радиатора отопления | 500 |
Установка закладных деталей в конструкции из ГКЛ | 450 |
Обшивка откосов ГКЛ в 1 слой до 300 мм | 350 |
Обшивка откосов ГКЛ в 1 слой 300-500 мм | 500 |
Обшивка откосов ГКЛ в 1 слой свыше 500 мм | 750 |
Устройство сантехнического короба из ГКЛ | 2000 |
Обшивка инсталляции ГКЛ | 1600 |
Устройство коробов из ГКЛ (1 слой ГКЛ) | 450 |
Устройство коробов из ГКЛ (2 слоя ГКЛ) | 650 |
Устройство коробов из ГКЛ для скрытой подсветки | 2000 |
Устройство ниш из ГКЛ под карнизы штор | 1500 |
Устройство простых ниш из ГКЛ (менее 1 кв. | 5000 |
Устройство простых ниш из ГКЛ (более 1 кв.м.) | 2500 |
Устройство сложных ниш из ГКЛ | 10000 |
Устройство ГКЛ перегородок (в 1 слой) | 800 |
Устройство ГКЛ перегородок (в 2 слоя) | 1000 |
Устройство ГКЛ перегородок (2 слоя усиленн.каркас) | 1800 |
Устройство арки из ГКЛ в 1 слой толщиной до 100 мм | 2000 |
Утепление и звукоизоляция |
|
|
|
 каркасу (в 1 слой)
каркасу (в 1 слой) м.)
м.)Устройство паро-теплоизоляции («пенофол», «пеноплекс») 420 р
Кладка из пазогребневых блоков или пеноблоков 450 р
Устройство пароизоляции («пенофол») 100 р
Устройство теплоизоляции («мин. вата»)
100 р
вата»)
100 р
Устройство теплоизоляции («пеноплекс») 320 р
Устройство звукоизоляции стен («Шуманет-БМ») 1 слой 150 р
Устройство звукоизоляции потолков («Аккустик Баттс») 200 р
Устройство звукоизоляции полов («Шуманет-100») 1 слой 180 р
Устройство звукоизоляции стен («Зипс Панель») с обшивкой в 1 слой ГКЛ) 650 р
Отделка потолка
Оштукатуривание потолков (по маякам) толщиной до 10 мм 420 р
Оштукатуривание потолков (по маякам) толщиной до 20 мм 660 р
Увеличение толщины штукатурки потолков на каждые 10 мм увеличения слоя 240 р
Выравнивание потолков (под правило) до 10 мм 300 р
Заделка потолочных (межпанельных) швов (рустов) 220 р
Заделка потолочных стыков ГКЛ (расшивка и шпаклевка) 240 р
Шпатлёвка потолков под покраску (комплекс работ: грунтовка, шпаклевка, шлифовка, армировка паутинкой или флизелином, финишная шпаклевка) 630 р
Окраска потолков в/э краской за 2 раза (с грунтовкой) 240 р
Обшивка потолков вагонкой, панелями (ПВХ,МДФ) 650 р
Устройство потолков типа Армстронг 400 р
Устройство потолков типа Армстронг (сложных) 650 р
Устройство реечных потолков 1000 р
Устройство натяжных потолков (пленка ПВХ) (комплекс) 700 р
Устройство натяжных потолков (ткань ПВХ) (комплекс) 1000 р
Установка потолочных карнизов (молдингов) 20-60 мм из полиуретана 240 р
Окраска потолочных карнизов (молдингов) 20-60 мм в/э краской за 2 раза 120 р
Окраска потолочных карнизов (молдингов) 60-120 мм в/э краской за 2 раза 180 р
Установка потолочных карнизов (молдингов) 70-120 мм из полиуретана 360 р
Окраска потолочных карнизов (молдингов) 60-120 мм в/э краской за 2 раза 180 р
Установка потолочных карнизов (молдингов) 130-200 мм из полиуретана 540 р
Окраска потолочных карнизов (молдингов) 130-200 мм в/э краской за 2 раза 270 р
Установка декоров из полиуретана 280 р
Отделка стен
Оштукатуривание стен (по маякам) толщиной до 10 мм 350 р
Оштукатуривание стен (по маякам) толщиной до 20 мм 550 р
Увеличение толщины штукатурки стен на каждые 10 мм увеличения слоя 200 р
Выравнивание стен штукатуркой «под правило» 250 р
Армирование штукатурки стен пластиковой сеткой 100 р
Монтаж металлической сетки под штукатурку 200 р
Заделка стеновых стыков ГКЛ (расшивка и шпаклевка) 200 р
Шпатлёвка стен под покраску (комплекс работ: грунтовка, шпаклевка, шлифовка, армировка паутинкой или флизелином, финишная шпаклевка) 500 р
Окраска стен в/э краской за 2 раза (с грунтовкой) 200 р
Шпатлёвка стен под обои (комплекс работ: грунтовка, шпаклевка, шлифовка) 250 р
Армирование стен малярной сеткой 120 р
Финишная шпаклевка стен под тонкие обои 200 р
Поклейка обоев (винил, флизелин) на стены без подбора рисунка 350 р
Поклейка обоев (винил, флизелин) на стены с подбором рисунка 500 р
Поклейка тонких обоев на стены без подбора 450 р
Поклейка тонких обоев на стены с подбором рисунка 700 р
Поклейка бумажных обоев на стены 450 р
Поклейка натуральных обоев (бамбук, джут и т. п.) на стены
600 р
п.) на стены
600 р
Шпатлёвка стен под фотообои (комплекс работ: грунтовка, шпаклевка, шлифовка, финишная шпаклевка) 380 р
Поклейка обоев под окраску 220 р
Покраска обоев в/э краской за 2 раза 200 р
Поклейка фотообоев на стены 500 р
Поклейка декоративного панно (фрески) на стены 1000 р
Поклейка рельефных обоев («дуплекс») 260 р
Поклейка текстильных обоев 700 р
Поклейка пробковых обоев 500 р
Нанесение жидких обоев на стены 600 р
Нанесение покрытия типа «Байрамикс» 750 р
Нанесение покрытия типа «Венецианская штукатурка» 1500 р
Обшивка стен вагонкой, панелями (ПВХ,МДФ) 650 р
Облицовка стен плиткой (стандартного размера 150-300 мм)
(простая укладка без подбора, бордюров и т. п.)
1200 р
п.)
1200 р
Облицовка стен крупноформатной плиткой (свыше 300 мм) 1380 р
Облицовка стен плиткой (стандартного размера 150-300 мм) с подбором рисунка 1440 р
Облицовка стен плиткой крупноформатной плиткой (свыше 300 мм) с подбором рисунка 1600 р
Облицовка стен мелкоформатной плиткой (до 150 мм) 1800 р
Облицовка стен «диким камнем», «клинкером» 1800 р
Облицовка стен мозаикой на матрице 2500 р
Облицовка стен мозаикой на матрице с подбором рисунка 3500 р
Облицовка стен мрамором, гранитом (толщиной до 12 мм) 2400 р
Сверление отверстий в плитке (диаметром до 120 мм) 300 р
Затирка швов керамической плитки однокомпонентной затиркой (моноколор) 200 р
Затирка швов керамической плитки двухкомпонентной затиркой (моноколор) 400 р
Затирка швов керамической плитки однокомпонентной затиркой(поликолор) 400 р
Затирка швов керамической плитки двухкомпонентной затиркой (поликолор) 800 р
Затирка швов мозаичной плитки (моноколор) 400 р
Затирка швов мозаичной плитки (поликолор) 800 р
Затирка швов «дикого камня» 300 р
Устройство полов
Устройство гидроизоляции полов (обмазочным способом) 300 р
Устройство полов из керамзитобетона (до 50 мм) 300 р
Увеличение толщины полов из керамзитобетона на каждые 10 мм 80 р
Устройство цементно-песчаной стяжки толщ. до 50 мм
350 р
до 50 мм
350 р
Увеличение толщины стяжки на каждые 10 мм 100 р
Установка армирующей металлической сетки 200 р
Армирование стяжки фиброволокном 100 р
Устройство нивелирующего наливого пола 2-3 мм 220 р
Устройство нивелирующего наливного пола 3-10 мм 250 р
Устройство нивелирующего наливного пола 10-30 мм 300 р
Устройство нивелирующего наливного пола 30-50 мм 350 р
Устройство чернового пола по деревянным лагам 650 р
Укладка керамзита (до 100 мм) 200 р
Монтаж фальшполов «Кнауф» (до 100 мм) 750 р
Монтаж фальшполов «Кнауф» более 100 мм (на каждые 10 мм увеличения толщины) 100 р
Настил на пол пленки ПВХ 30 р
Укладка ГВЛВ (1 слой) 160 р
Укладка фанеры калиброванной (1 слой) 200 р
Шлифовка фанеры 100 р
Укладка ламината с подложкой (прямая укладка) 400 р
Укладка ламината с подложкой (диагональная укладка) 460 р
Укладка паркетной доски ( в «замок») (с подложкой) 400 р
Укладка паркетной доски ( в «замок») по диагонали 460 р
Укладка паркетной доски (массива) на клей 800 р
Укладка паркетной доски (массива) на клей по диагонали 950 р
Дополнительное крепление паркетной доски саморезами 200 р
Укладка кварцвинила ( в «замок») (прямая укладка) 350 р
Укладка кварцвинила ( в «замок») (диагональная укладка) 400 р
Укладка пробкового покрытия на клей 800 р
Покрытие полов лаком (1 слой) 100 р
Укладка штучного паркета (комплекс работ: укладка, циклевка, лакировка в 3 слоя) 1500 р
Настил рулонного линолеума 250 р
Настил коммерческого линолеума 300 р
Проварка швов линолеума 100 р
Настил рулонного коврового покрытия 250 р
Настил плиточного коврового покрытия 400 р
Фигурная резка ламината, паркетной доски 350 р
Укладка плитки на пол (размером 150-300 мм) 1000 р
Укладка плитки на пол (размером 150-300 мм) по диагонали 1200 р
Укладка плитки крупного формата (свыше 300 мм) на пол 1150 р
Укладка плитки на пол с подбором рисунка 1200 р
Укладка плитки нестандартного размера на пол с подбором рисунка 1350 р
Укладка мозаики (на матрице) на пол 2000 р
Укладка мозаики (на матрице) на пол с подбором рисунка 2400 р
Укладка мрамора на пол 2000 р
Затирка швов керамической плитки однокомпонентной затиркой (моноколор) 200 р
Затирка швов керамической плитки двухкомпонентной затиркой (моноколор) 400 р
Затирка швов керамической плитки однокомпонентной затиркой(поликолор) 400 р
Затирка швов керамической плитки двухкомпонентной затиркой (поликолор) 800 р
Затирка швов мозаичной плитки (моноколор) 400 р
Затирка швов мозаичной плитки (поликолор) 800 р
Установка накладного порожка 450 р
Установка пробкового компенсатора 600 р
Установка плинтуса (пластик) 220 р
Установка плинтуса (массив, шпон, МДФ) до 60 мм. 300 р
300 р
Установка плинтуса (массив, шпон, МДФ) 60-100 мм 360 р
Установка плинтуса (массив, шпон, МДФ) выше 100 мм. 400 р
Устройство плинтусов из плитки (готовый плинтус) 250 р
Устройство плинтусов из плитки (с изготовлением) 400 р
Окраска плинтуса до 60 мм. 200 р
Окраска плинтуса 60-100 мм. 250 р
Окраска плинтуса свыше 100 мм. 300 р
300 р
Установка плинтуса из пробки 450 р
Облицовка ступеней плиткой 2000 р
Облицовка порогов плиткой 2500 р
Облицовка порогов мозаикой 5000 р
Другие отделочные работы
Оштукатуривание откосов до 100 мм 250 р
Оштукатуривание откосов 100-200 мм 350 р
Оштукатуривание откосов свыше 200 мм 450 р
Оштукатуривание откосов арочных до 100 мм 600 р
Облицовка откосов плиткой 1000 р
Облицовка откосов мозаикой 2000 р
Облицовка откосов мрамором 1200 р
Обшивка откосов вагонкой, панелями (ПВХ,МДФ) 500 р
Шпатлёвка откосов под окраску (комплекс работ: грунтовка, шпаклевка, шлифовка,армировка паутинкой или флизелином, финишная шпаклевка) 350 р
Окраска откосов в/э краской за 2 раза (с грунтовкой) 150 р
Оштукатуривание колонн (сторона до 100 мм) 1000 р
Оштукатуривание колонн (сторона 100-200 мм) 2000 р
Оштукатуривание колонн (сторона свыше 200 мм) 1600 р
Шпатлёвка коробов (сторона до 100 мм) под окраску (комплекс работ) 480 р
Окраска коробов (сторона до 100 мм) в/э краской за 2 раза 220 р
Шпатлёвка коробов (сторона 100-200 мм) под окраску (комплекс работ) 680 р
Окраска коробов (сторона 100-200 мм) в/э краской за 2 раза 300 р
Шпатлёвка коробов (сторона свыше 200 мм) под окраску (комплекс работ) 960 р
Окраска коробов (сторона свыше 200 мм) в/э краской за 2 раза 440 р
Шпаклевка коробов из ГКЛ для скрытой подсветки (комплекс работ) 950 р
Окраска коробов из ГКЛ для скрытой подсветки в/э краской за 2 раза 440 р
Шпаклевка ниш из ГКЛ под карнизы штор (комплекс работ) 650 р
Окраска ниш из ГКЛ под карнизы штор в/э краской за 2 раза 400 р
Установка малярных уголков 60 р
Установка арочных уголков на сложные поверхности 120 р
Установка пластиковых подоконников до 300 мм 600 р
Установка пластиковых откосов до 300 мм 500 р
Установка пластиковых уголков 200 р
Окраска дверных и оконных блоков 1200 р
Окраска радиаторов 650 р
Окраска труб диаметром до 50 мм 120 р
Электромонтажные работы
Устройство временного электроснабжения 40 р
Диагностика электрической сети 1200 р
Штробление до 30х30 мм 400 р
Заделка штроб до 30х30 мм 100 р
Прокладка кабеля (силового и слаботочного) 80 р
Установка подрозетника (с устройством гнезда) 400 р
Перенос подрозетника (с устройством гнезда) (на расстояние. до 2-х метров) 1000
р
до 2-х метров) 1000
р
Установка подрозетника — распайки (с устройством гнезда и подключением) 750 р
Установка накладной распаечной коробки (с подключением) 500 р
Установка распределительного эл.щита накладного 1000 р
Установка распределительного эл.щита встраиваемого (12 модулей) 2400 р
Установка распределительного эл.щита встраиваемого (24 модулей) 3600 р
Установка распределительного эл.щита встраиваемого (36 модулей) 4800 р
Установка распределительного эл.щита встраиваемого (48 модулей) 5400 р
Установка распределительного эл.щита встраиваемого (60 модулей) 8000 р
Установка автоматов защиты (1 полюсных) 150 р
Установка УЗО, дифавтоматов, автоматов (2-х полюсных) 300 р
Установка УЗО, дифавтоматов, автоматов (3-х и 4-х полюсных) 500 р
Установка электросчетчика (без пломбировки) 1500 р
Установка розетки, выключателя, диммера 300 р
Установка проходного выключателя 500 р
Установка TВ, Tелефонной, Интернет. USB розетки
400 р
USB розетки
400 р
Перенос розетки, выключателя (на расст. до 2-х метров) 2000 р
Установка TВ разветвителя (краба) 500 р
Установка интернет-роутера (с подключением) 1500 р
Установка люстры (2-5 рожков) (без сборки) 1000 р
Установка подвесного (накладного) светильника 300 р
Установка встроенного (точечного) светильника 400 р
Установка подвесного (трекового) светильника 500 р
Установка встроенного (линейного) светильника 2000 р
Установка бра 300 р
Установка светодиодной ленты 200 р
Установка трансформатора 450 р
Устройство электровыводов 100 р
Подключение подсветок зеркал, шкафов и т. д.
150 р
д.
150 р
Установка звонка 500 р
Установка аудиодомофона 600 р
Установка видеодомофона 3000 р
Установка электрического полотенцесушителя 1000 р
Устройство электрических теплых полов 500 р
Установка регулятора теплых полов 500 р
Подключение водонагревателя 500 р
Подключение ввода 220 вольт 2200 р
Подключение ввода 380 вольт 3300 р
Подключение TВ, Tелефонного, Интернет ввода 1000 р
Установка сушилки для рук 300 р
Установка металлических лотков под проводку 200 р
Монтаж кабель-канала под проводку 150 р
Разметка мест установки электрики (без проекта) 100 р
Сантехнические работы
Устройство временного водоснабжения 1000 р
Устройство сантехнической штробЫ 600 р
Заделка сантехнической штробЫ 200 р
Установка шарового крана 350 р
Установка фильтра грубой очистки 350 р
Установка счетчика воды (без пломбировки) 1450 р
Установка регулятора давления (редуктора) 450 р
Установка фильтра тонкой очистки воды с регулятором давления, манометром и прямоточной промывкой 1500 р
Установка фильтра очистки питьевой воды (типа Аквафор, Гейзер, Барьер, Атолл) «под ключ»2500
Установка обратного клапана 350 р
Установка коллектора на 2-8 выходов (комплекс работ) 2500 р
Установка водорозетки 500 р
Прокладка труб ХГВ (cшитый полиэтилен РЕХАУ) 300 р
Прокладка канализационных труб (ПВХ) 500 р
Тепло-звукоизоляция труб 100 р
Опрессовка системы водоснабжения (1 стояк) 1200 р
Врезка в канализационный стояк 2200 р
Установка накопительного водонагревателя (бойлера) 3500 р
Установка проточного водонагревателя 2500 р
Установка инсталляции 4200 р
Установка подвесного (приставного) унитаза 2500 р
Установка клавиши слива инсталляциИ 500 р
Установка унитаза «Компакт» 2800 р
Установка биде 2500 р
Установка писсуара 2400 р
Установка раковины 2000 р
Установка раковины с тумбой 3200 р
Установка смесителя 850 р
Установка смесителя скрытого монтажа 2000 р
Установка гигиенического душа 1000 р
Установка встроенного гигиенического душа 2500 р
Установка водяного полотенцесушителя (с подводкой труб) 3500 р
Установка водяного полотенцесушителя на готовое место 1500 р
Подключение стиральной машины 1000 р
Установка ванны простой (сталь, чугун, акрил) 4000 р
Установка смесителя ванны (душевого гарнитура) 1200 р
Установка верхнего душа (тропический душ) 2200 р
Установка готового экрана ванны (ПВХ, МДФ) 1000 р
Установка гидромассажной ванны 8500 р
Установка душевого бокса 150-170 см 9000 р
Установка душевой кабины с поддоном 7600 р
Установка трапа (лотка) душевой кабины 1200 р
Установка душевой панели 3000 р
Установка поддона душевой кабины 2000 р
Облицовка поддона душевой кабины мозаикой (на матрице) 3000 р
Установка дверок душевой кабины 4500 р
Подключение посудомоечной машины 2400 р
Установка системы контроля протечки воды (2 крана + 1 контроллер) с беспроводными 2-4 датчиками (без штробления и прокладки проводов) 6500 р
Установка системы контроля протечки воды (2 крана + 1 контроллер) с проводными 2-4 датчиками (без штробления и прокладки проводов) 8500 р
Монтаж системы отопления
Замена радиатора отопления с переделкой узла подводки (комплекс работ) 7500 р
Замена стояка отопления 5000 р
Переварка узла подводки радиатора отопления 4500 р
Переделка узла подводки радиатора отопления 4500 р
Прокладка труб отопления (сшитый полиэтилен REXAU) 300 р
Теплоизоляция труб отопления 100 р
Врезка в водопроводный стояк 4500 р
Установка шарового крана, терморегулятора 300 р
Установка радиатора отопления на готовое место 1500 р
Установка встроенного конвектора 10000 р
Установка накладного шкафа коллектора отопления 1000 р
Установка встроенного шкафа коллектора отопления 5000 р
Установка коллектора отопления (на 2-8 выходов) 2800 р
Переделка узла подводки полотенцесушителя 12000 р
Снятие и установка радиатора для проведения отделочных работ за радиатором (3-4 раза) 2000 р
Вентиляционные работы
Врезка в вентиляционный короб 1000 р
Монтаж вентиляционного канала (до 2 м) 2000 р
Установка вентилятора (с подключением) 500 р
Установка вентиляционной решетки 220 р
Установка дверей
Установка одностворчатой двери с врезанной фурнитурой 3500 р
Установка двустворчатой двери с врезанной фурнитурой 5000 р
Установка раздвижной двери типа «купе» (без установки пенала) 3500 р
Установка готового пенала для раздвижной двери типа «купе»
5000 р
Установка двери типа «Гармошка» 2000 р
Врезка петель 200 р
Врезка ручки-защёлки 300 р
Установка наличников 100 р
Установка доборов (за каждые 10 см. ширины)
120 р
ширины)
120 р
Изготовление доборов 100 р
Врезка дверного замка 550 р
Врезка «глазка» 250 р
Установка декоративных стекол 220 р
Установка отбойника 220 р
Установка доводчика 300 р
Демонтажные работы
Демонтаж перегородок из кирпича (1/2 кирпича) 320 р
Резка перегородок из кирпича (1/2 кирпича) 500 р
Устройство проема в кирпичной стене (1/2 кирпича) 1300 р
Демонтаж перегородок из кирпича (1 кирпич) 640 р
Резка перегородок из кирпича (1 кирпич) 1000 р
Устройство проема в кирпичной стене (1 кирпич) 2500 р
Демонтаж перегородок из блоков до 100 мм 220 р
Резка перегородок из блоков толщ. до 100 мм
400 р
до 100 мм
400 р
Устройство проема в блочной стене толщ. до 100 мм 1000 р
Демонтаж перегородок из бетона, ж/б до 100 мм 1000 р
Демонтаж перегородок из бетона, ж/б до 200 мм 2000 р
Резка перегородок из бетона, ж/б до 100 мм 1250 р
Резка перегородок из бетона, ж/б до 200 мм 2500 р
Резка перегородок из бетона, ж/б до 300 мм 3700 р
Устройство проема в бетоне,ж/б толщ. до 100 мм 3200 р
Демонтаж сантехкабины из ацеита (стены и потолок) 300 р
Демонтаж поддона сантехкабины из бетона 1250 р
Демонтаж перегородок из ацеита 250 р
Демонтаж перегородок из ГКЛ 250 р
Демонтаж перегородок из дерева, ДСП толщ. до 100 мм
300 р
до 100 мм
300 р
Демонтаж подоконной части стены из ПГП, пенобетона 650 р
Демонтаж подоконной части стены из кирпича 1300 р
Демонтаж подоконной части стены из бетона, ж/б 4800 р
Устройство отверстий в бетонных стенах (до 20 мм) 100 р
Устройство отверстий в бетонных стенах (до 50 мм) 250 р
Устройство отверстий в бетонных стенах (до 100 мм) 500 р
Снятие полов из досок (с лагами) 200 р
Снятие полов из щитового паркета (с лагами) 280 р
Снятие сухой засыпки полов (до 100 мм) 150 р
Снятие паркетных полов (на клею, мастике) 250 р
Снятие ламината, паркетой доски 100 р
Снятие фанеры 1 слой 120 р
Снятие оргалита (на мастике) 250 р
Снятие линолеума, ковролина 100 р
Демонтаж плинтусов 50 р
Демонтаж плитки, керамогранитаь 120 р
Демонтаж плитки, керамогранита (особой прочности) 250 р
Демонтаж мрамора, гранита 220 р
Снятие пластиковых уголков 50 р
Расшивка потолочных швов (рустов) 150 р
Удаление в/э краски (побелки),шпаклевки с потолков 200 р
Частичное удаление в/э краски (побелки) с потолков 100 р
Удаление в/э краски (побелки),шпаклевки со стен 150 р
Удаление масляной краски, шпаклевки с потолков 250 р
Удаление масляной краски, шпаклевки со стен 200 р
Удаление обоев (1 слой), шпаклевки со стен 120 р
Удаление обоев (1 слой), шпаклевки с потолков 150 р
Снятие штукатурки толщ. до 30 мм со стен 180
р
до 30 мм со стен 180
р
Снятие штукатурки толщ. до 60 мм со стен 350 р
Снятие штукатурки до 30 мм(особо прочной) со стен 350 р
Снятие штукатурки толщ. до 30 мм с потолков 200 р
Снятие штукатурки толщ. до 60 мм с потолков 420 р
Снятие штукатурки толщ.до 30 мм (особо прочной) с потолков 420 р
Демонтаж керамзитобетонных полов (до 50 мм) 120 р
Демонтаж керамзитобетонных полов (до 100 мм) 240 р
Снятие цементно-песч. стяжки ( до 50 мм ) 180
р
стяжки ( до 50 мм ) 180
р
Снятие цементно-песч. стяжки ( до 100 мм ) 360 р
Демонтаж бетонных полов (до 50 мм) армиров. 1250 р
Демонтаж бетонных полов (до 100 мм) армиров. 2500 р
Демонтаж обшивки стен ГКЛ (1 слой) ( с каркасом) 180 р
Демонтаж сухой штукатурки со стен (1 слой) 120 р
Демонтаж обшивки стен и потолков из вагонки и панелей ПВХ (с каркасом) 150 р
Демонтаж клееных панелей ПВХ на стенах и потолке 100 р
Демонтаж антресолей, встроенных шкафов, ниш и т. п.
180 р
п.
180 р
Демонтаж подвесных потолков «Армстронг» 100 р
Демонтаж реечных потолков 100 р
Демонтаж натяжных потолков 60 р
Демонтаж потолков ГКЛ (с каркасом) 240 р
Демонтаж оконных блоков 250 р
Демонтаж подоконников из дерева или ПВХ 100 р
Демонтаж бетонных подоконников 250 р
Демонтаж дверного блока 350 р
Демонтаж дверного блока из стали 1000 р
Демонтаж электропроводки (площадь помещений) 30 р
Демонтаж розеток, выключателей, светильников 90 р
Демонтаж подрозетника с заделкой гнезда 150 р
Демонтаж электрощита в сборе 500 р
Демонтаж электроплиты 200 р
Демонтаж электрического полотенцесушителя 150 р
Демонтаж кранов 110 р
Демонтаж фильтра грубой очистки 100 р
Демонтаж счетчика воды 200 р
Демонтаж фильтра тонкой очистки 350 р
Демонтаж регулятора давления 220 р
Демонтаж коллектора 500 р
Демонтаж сантехнических труб 120 р
Демонтаж смесителя 220 р
Демонтаж ванны (сталь, акрил) 1000 р
Демонтаж ванны (чугун) 1500 р
Демонтаж унитаза, биде 400 р
Демонтаж раковины «Тюльпан» 300 р
Демонтаж раковины с тумбой 500 р
Демонтаж водяного полотенцесушителя 650 р
Демонтаж водонагревателя 450 р
Демонтаж гидромассажной ванны 1200 р
Демонтаж душевой кабины 1000 р
Демонтаж стиральной машины 250 р
Демонтаж инсталляции 500 р
Демонтаж радиатора отопления 500 р
Демонтаж труб отопления 150 р
Демонтаж газовой плиты 500 р
Прочие работы
Вынос мусора (контейнер 8 м3) (на грузовом лифте) 5000 р
Вынос мусора (контейнер 8 м3) (на негрузовом лифте) 6000 р
Вынос мусора (в местный контейнер) (на грузовом лифте) 80 р
Вынос мусора (в местный контейнер) (на негрузовом лифте) 100 р
Вынос мусора (в местный контейнер) (без лифта) за 1 этаж 80 р
Рагрузка и подъем материалов (на грузовом лифте) 1200 р
Рагрузка и подъем материалов (на негрузовом лифте) 1500 р
Рагрузка и подъем материалов (без лифта) за 1 этаж 1200 р
Укрытие полов пленкой ПВХ 30 р
Укрытие полов оргалитом 60 р
Где и на кого учиться, чтобы стать предпринимателем
Многие задаются вопросом, где и на кого учиться, чтобы стать предпринимателем? Что же, на самом деле, предпринимателем, конечно, становятся по факту предпринимательской деятельности. Но есть несколько способов, получить знания, которые помогут вам стать успешным предпринимателем.
Но есть несколько способов, получить знания, которые помогут вам стать успешным предпринимателем.
Вы можете получить эти знания из книг, тренингов, курсов, бесплатных видео на Youtube и даже формальных образовательных программ. В этой статье мы расскажем обо всех возможностях, а также расскажем, знания из каких сфер нужны предпринимателю.
Чем старше становлюсь, тем больше чувствую потребность в обучении. Все сферы жизни развиваются с огромной скоростью и чтобы быть не только вкурсе, но и не отстать, сегодня нам нужно постоянно повышать квалификацию и приобретать новые навыки. Хорошо что есть все возможные формы обучения, но в противовес и все возможные формы развлечения, которые отвлекают от обучения (простите за тавтологию). ✅Итак один из главных пунктов в моем плане на год это саморазвитие и обучение, кроме книг и множества видео материала я посещаю тренинги и семинары, но их стало так много, а толковых мало, большинство стараются либо собрать денег, либо хайпануть, очень мало толковых учителей, тренеров и мероприятий.
Публикация от ⠀⠀⠀⠀⠀ ⠀⠀⠀⠀Костя Пекинец (@kostya_pekinec)
 Один из признанных мировых лидеров в развитии личности – Тони Роббинс, если честно он лучший И я скоро еду на его программу «Свидание с судьбой» в Сингапур. Ещё важная мысль которую я понял в обучении-за обучение нужно платить, если не платим, не ценим! Я знаю что многие скептически настроены к такому обучению, это ваше право, но есть и те кто поняли хорошее влияние правильных людей! Поэтому спрошу, какие тренера, коучи, учителя повлияли на вас? Просьба не пишите мама и папа, я о другом. #тренер #коучинг #коуч #тренинги #семинар #тонироббинс #сингапур
Один из признанных мировых лидеров в развитии личности – Тони Роббинс, если честно он лучший И я скоро еду на его программу «Свидание с судьбой» в Сингапур. Ещё важная мысль которую я понял в обучении-за обучение нужно платить, если не платим, не ценим! Я знаю что многие скептически настроены к такому обучению, это ваше право, но есть и те кто поняли хорошее влияние правильных людей! Поэтому спрошу, какие тренера, коучи, учителя повлияли на вас? Просьба не пишите мама и папа, я о другом. #тренер #коучинг #коуч #тренинги #семинар #тонироббинс #сингапурПредпринимателю нужно учиться бизнесу в целом, маркетингу и продажам, нужно прокачивать навыки саморазвития и мышления, а также получать знания из той отрасли, в которой он намерен работать.
Я не буду давать, конечно, исчерпывающий перечень материалов, но предложу довольно много источников для старта.
Уже готовы действовать? Тогда присоединяйтесь к двум важнейшим курсам по бизнесу: крупнейшая в России онлайн-конференция по созданию своего бизнеса «Концентрат» и онлайн-марафон «Ты – предприниматель». Вы научитесь создавать бизнес в реальных условиях: работа, учёба, кредиты и ипотека, а также другие обязательства. Сделаете шаг к свободе и финансовому благополучию. Вашими наставниками будут реальные предприниматели, а не преподаватели-теоретики.
Посмотреть эту публикацию в Instagram
🚀Антикризисный «Концентрат» стартует уже сегодня.
Публикация от LIKE Центр Пермь (@likebz59)
 ⠀ А ниже 17 причин, почему стоит быть сегодня с нами: ⠀ 1. Получишь список прибыльных ниш и эффективных каналов трафика. 2. Научишься отличать якобы «перспективную идею» от пуленепробиваемого факта. 3. Поймешь, как вернуть старых клиентов и привлечь новых, не спуская последнюю рубаху на маркетинговый бюджет. 4. Узнаешь, как и где искать подрядчиков, клиентов, инвесторов и суперпрофессионалов в команду (спойлер: их можно найти прямо на «Концентрате»). 5. Научишься отрабатывать возражения клиентов, для которых каждая копейка на счету. 6. Узнаешь, что предложить клиенту в кризис, чтобы даже самый дотошный и упрямый человек согласился. 7. Попрактикуешься продавать прямо на интенсиве, не вставая с дивана. 8. Поймешь, в какой проект стоит инвестировать, пока все складируют накопления в маски и гречку. 9. Узнаешь, как сохранить команду, даже если есть упаднические настроения и проблемы с зарплатой. 10. Разберешься, что делать, если работа не нравится, а деньги нужны. И на чем ты можешь заработать уже сейчас.
⠀ А ниже 17 причин, почему стоит быть сегодня с нами: ⠀ 1. Получишь список прибыльных ниш и эффективных каналов трафика. 2. Научишься отличать якобы «перспективную идею» от пуленепробиваемого факта. 3. Поймешь, как вернуть старых клиентов и привлечь новых, не спуская последнюю рубаху на маркетинговый бюджет. 4. Узнаешь, как и где искать подрядчиков, клиентов, инвесторов и суперпрофессионалов в команду (спойлер: их можно найти прямо на «Концентрате»). 5. Научишься отрабатывать возражения клиентов, для которых каждая копейка на счету. 6. Узнаешь, что предложить клиенту в кризис, чтобы даже самый дотошный и упрямый человек согласился. 7. Попрактикуешься продавать прямо на интенсиве, не вставая с дивана. 8. Поймешь, в какой проект стоит инвестировать, пока все складируют накопления в маски и гречку. 9. Узнаешь, как сохранить команду, даже если есть упаднические настроения и проблемы с зарплатой. 10. Разберешься, что делать, если работа не нравится, а деньги нужны. И на чем ты можешь заработать уже сейчас. 11. Получишь пошаговый план для декомпозиции финансовых целей. Поймешь, как перестать мечтать о миллионе и начать потихоньку его зарабатывать. 12. Научишься экономить сбережения, оптимизировать и реструктуризировать долги. И забудешь фразу «одолжи косарь по-братски» как страшный сон. 13. Узнаешь, как оцифровать бизнес и обеспечить финансовый резерв. 14. Получишь инструкцию по переводу бизнеса в онлайн и список ниш, в которые стоит зайти прямо сейчас. 15. Разберешься в кредитных каникулах, льготах и мерах господдержки. Ведь ты можешь рассчитывать на хорошие деньги и не знать об этом. 16. Заведешь полезные и приятные знакомства. 17. Получишь заряд бодрости и мотивации, которые помогут выехать из самоизоляции на коне. ⠀ ❗И последняя, самая важная причина: ты получишь всё это великолепие за 500 рублей (по цене нескольких масок, которые всё равно не помогут, если кто-то чихнет на тебя в лифте). ⠀ Успей зарегистрироваться – проведи выходные с пользой. ⠀ #likebz #likebz #likebz59 #концентрат #likefamilyperm #бизнеспермь #пермьактивная
11. Получишь пошаговый план для декомпозиции финансовых целей. Поймешь, как перестать мечтать о миллионе и начать потихоньку его зарабатывать. 12. Научишься экономить сбережения, оптимизировать и реструктуризировать долги. И забудешь фразу «одолжи косарь по-братски» как страшный сон. 13. Узнаешь, как оцифровать бизнес и обеспечить финансовый резерв. 14. Получишь инструкцию по переводу бизнеса в онлайн и список ниш, в которые стоит зайти прямо сейчас. 15. Разберешься в кредитных каникулах, льготах и мерах господдержки. Ведь ты можешь рассчитывать на хорошие деньги и не знать об этом. 16. Заведешь полезные и приятные знакомства. 17. Получишь заряд бодрости и мотивации, которые помогут выехать из самоизоляции на коне. ⠀ ❗И последняя, самая важная причина: ты получишь всё это великолепие за 500 рублей (по цене нескольких масок, которые всё равно не помогут, если кто-то чихнет на тебя в лифте). ⠀ Успей зарегистрироваться – проведи выходные с пользой. ⠀ #likebz #likebz #likebz59 #концентрат #likefamilyperm #бизнеспермь #пермьактивнаяСодержание статьи
Бизнес
В принципе, научиться вести бизнес – это то, что можно сделать даже в стенах Вуза. Я дам ссылки как на практические, так и теоретические источники.
Я дам ссылки как на практические, так и теоретические источники.
Другое дело, что помимо знаний собственно о бизнесе, вам потребуется много чего еще, чему в университетах не учат, да и не могу научить. Но об этом ниже.
Книги о бизнесе
Самая известная в мире образовательная программа о бизнесе называется MBA – Master of Business Administration, то есть Магистр делового администрирования. Этой программе учат как обычные университеты, так и специализированные бизнес-школы (получить MBA в крутой бизнес-школе считается наиболее престижным вариантом).
Есть много книг, которые знакомят с ключевыми принципами программ MBA, вот несколько хороших:
Классические книги, которые читает большинство студентов бизнес-школ:
Несколько полезных и важных книг о бизнесе:
Книги про старт бизнеса:
Книги, написанные успешными предпринимателями:
Бизнес-знания на Youtube
Подпишитесь на канал университета Синергия, канал вебинаров от университета Синергия и канал тренинговой компании Бизнес-Молодость.
И я вам сразу предложу несколько видео для начала.
Выступление молодого миллионера из Ижевска Аяза Шабутдинова, его опыт в бизнесе и много чего еще интересного.
Выступление бизнес-тренера из Украины Ицхака Пинтосевича на актуальную для всех жителей России тему, как стать миллиардером.
Бизнес-дзен от предпринимателя и Интернет-маркетолога Руслана Татунашвили. Вообще, это видео посмотрите сами и рекомендуйте тем людям, которые находятся в трудной жизненной ситуации, которым сейчас тяжело.
Предприниматель Артем Агабеков, какие советы он бы дал себе 20-летнему (полезно не только 20-летним).
Статьи
Несколько полезных статей на нашем сайте:
Курсы онлайн и офлайн
Образовательные курсы от компании Бизнес-Молодость, прекрасный вариант, если вы уже все посмотрели, все почитали, что-то начали, но все идет не так, как вам хотелось бы. Для совсем зеленого новичка больше подойдут бесплатные видео на Youtube-канале Бизнес-Молодость, к счастью, ребята очень охотно делятся качественным контентом, на Youtube можно найти массу полезных семинаров от БМ, некоторые длятся много часов.
Для совсем зеленого новичка больше подойдут бесплатные видео на Youtube-канале Бизнес-Молодость, к счастью, ребята очень охотно делятся качественным контентом, на Youtube можно найти массу полезных семинаров от БМ, некоторые длятся много часов.
Like Центр – это более молодая команда и очень-очень сильный конкурент Бизнес-Молодости. Продукты у них похожие, но у Like есть уникальный трехдневный формат, который называется Концентрат. Стоит он очень недорого, длится три дня, а не 2 месяца. И для многих предпринимателей Концентрата уже хватает, чтобы запустить бизнес и заработать первые деньги.
Курсы на сервисе Открытое образование, к сожалению, по бизнесу там совсем небольшой выбор.
UNIWEB – у них больше курсов по бизнесу, многие можно попробовать бесплатно.
Универсариум – тоже довольно много курсов по бизнесу, некоторые лекции выложены бесплатно.
Университет Синергия также предлагает несколько онлайн-курсов, каждый курс включает в себя несколько занятий и обычно дает возможность живого общения с преподавателем.
Образовательные учреждения
В Университете Синергия есть курс Предпринимательства (первое высшее образование) и много других образовательных курсов. Обучение в университете платное, для старта в бизнесе, наверное, это сегодня лучшее учебное заведение.
Высшая школа экономики имеет филиалы во многих городах России, обычно филиалы ВШЭ – это лучший вариант высшего образования в тех городах, в которых они представлены. У них есть качественные программы бакалавриата по Маркетингу и Управлению бизнесом.
Бизнес-школа Сколково – лучшая, пожалуй, бизнес-школа в стране. В Сколково могут учиться состоявшиеся люди, предприниматели и менеджеры. Сразу после школьной скамьи в Сколково не примут.
Маркетинг
Маркетинг – это ключевой навык, который пронизывает любой бизнес от начала до конца. Хороший маркетинг заключается в том, чтобы придумать, какой продукт нужно сделать, чтобы он превзошел существующие на рынке альтернативы и конкурентов, а также понять, как убедить потребителей приобрести этот продукт.
Прекрасное знание маркетинга сделает вас очень успешным, богатым и востребованным человеком.
Что касается курсов онлайн и образовательных учреждений, то рекомендации здесь и дальше такие же, что в разделе про бизнес. Но я предложу несколько полезных книг и бесплатных видео.
Книги
Классические книги по маркетингу:
Другие хорошие книги по маркетингу:
Маркетинговые знания на Youtube
Много маркетинговых видео вы найдете на тех каналах, которые я вам уже рекомендовал. Маркетинг – это основа любого бизнеса, так что не удивительно, что ему уделяется больше внимания, чем любой другой теме в бизнесе.
Если вы вообще ничего не знаете о маркетинге, вам обязательно нужно начать с книг, сразу смотреть видео будет сложновато.
Я сразу предлагаю вам несколько хороших видео про маркетинг.
Игорь Манн – не только ведущий российский эксперт по маркетингу, но и очень крутой эксперт на международном уровне.
 youtube.com/embed/9X-8JiOhzX4″ frameborder=»0″ allowfullscreen=»allowfullscreen»/>
youtube.com/embed/9X-8JiOhzX4″ frameborder=»0″ allowfullscreen=»allowfullscreen»/>Игорь Манн – автор многих книг, совладелец нескольких успешных компаний, он много выступает, в сети доступно большое количество его видео, причем не только про маркетинг.
Реальный маркетинг – это целая линейка образовательных продуктов от тренинговой компании Бизнес-Молодость, много видео из этих курсов выложено на Youtube.
Выступление Руслана Татунашвили для курса Реальный маркетинг.
Выступление Артема Агабекова про маркетинг.
Продажи
Плохие продажи могут погубить даже самый прекрасный бизнес с самой лучшей и продуманной маркетинговой стратегией. Хорошие продажи спасут даже самый проблемный бизнес с самой паршивой маркетинговой стратегией.
Хорошие продажи спасут даже самый проблемный бизнес с самой паршивой маркетинговой стратегией.
В общем, учитесь хорошо продавать, этот навык нужен абсолютно любому предпринимателю, да и пригодится любому человеку вообще.
Классные книги про продажи:
Несколько полезных видео про продажи на Yotube.
Ицхак Пинтосевич – прекрассный бизнес-тренер и автор нескольких книг, в том числе книги “Продавай!”.
Настасья Белочкина – ведущий российский эксперт по продажам (ну или не российский, вообще-то Настасья из Беларуси).
Настасья – руководитель конслатинговой компании, которая помогает другим компаниям продавать больше.
Саморазвитие
Предприниматель постоянно сталкивается со все более сложными задачами, рискует каждый день, отвечает не только за себя и свою семью, но и за многих других людей.
Карьера предпринимателя – это всегда тонкий баланс между полным крахом и полной победой. Предприниматели, которые не прокачивают навык саморазвития, рано или поздно перестают быть предпринимателями, так как терпят крушение, после которого уже не могут оправиться.
К счастью, сегодня саморазвитие – это вполне проработанная область знаний, есть много книг, видео и тренингов, которые вам помогут.
Полезные книги о саморазвитии, мотивации и самоорганизации:
Очень много классных видео выложено в бесплатном доступе на Youtube.
Подпишитесь на тренинговую компанию Business Relations. Они также проводят очень полезный тренинг Контекст (и другие классные тренинги тоже).
Обязательно посмотрите несколько видео Радислава Гандапаса.
Подпишитесь на его канал на Youtube.
И если будет возможность, сходите на его тренинг.
Посмотрите несколько видео Ицхака Пинтосевича и подпишитесь на канал его тренинговой компании.
И подпишитесь на русскоязычный Youtube-канал Тони Роббинса (у Тони Роббинса есть еще много книг, но читать их лично мне сложновато, слишком много “воды”, поэтому рекомендую слушать аудиокниги Тони Роббинса).
Несколько полезных статей на нашем сайте и на других сайтах:
Присоединяйся к марафону «Ты – предприниматель» от наших экспертов, опытных предпринимателей. На марафоне любой сможет создать или прокачать свой бизнес, сделав шаг к финансовому благополучию.
Посмотреть эту публикацию в Instagram
В 35 понял, что готов делиться своим опытом ⠀ Привет, мой друг👋🏻 Недавно мне исполнилось 35, и я понял, что за свои годы я уже открыл 5 бизнесов ⠀ Были успешные и неуспешные проекты. Я проходил несколько раз путь от многорукого многонога до отдельного кабинета. Это были сезонные и не зависящие от сезона варианты ⠀ Но я понял что у меня есть навык: 📍 Придумывать, запускать, хоронить и, конечно, развивать идеи 📍 Создавать бизнес 📍 Формировать команду 📍 Оцифровывать бизнес-процессы 📍 Анализировать ⠀ Я имею опыт запуска 5ти бизнесов в разных сферах: 1. Ремонт, обслуживание, продажа инструмента и бытовой техники 2. IT 3. ЖКХ 4. Изготовление и продажа рабочих ХБ перчаток 5. Маркетинговое агентство по изготовлению наружной рекламы и печатной продукции ⠀ И тем бэкграундом, тем опытом, который у меня есть, я готов поделиться с тобой 🙌🏻 ⠀ Готов проконсультировать тебя бесплатно: 👉🏻 Если ты начинающий предприниматель 👉🏻 Действующий предприниматель, но чувствуешь упадок сил ⠀ Пиши💥 будем решать твой вопрос😌
Публикация от Антон Стёпин (@stepin_mentor)
Мышление
Предприниматель может вообще ни в чем глубокого не разбираться и ничего особенно хорошо не уметь. Кроме одного важного навыка. Предприниматель должен уметь хорошо думать, находить нестандартные решения и придумывать новые идеи.
Кроме одного важного навыка. Предприниматель должен уметь хорошо думать, находить нестандартные решения и придумывать новые идеи.
Книги, которые научат вас думать лучше:
Сервис, который поможет вам думать – Writelight.
Ваша отрасль
И все же на старте, вероятно, вам придется очень хорошо разобраться в той отрасли, в которой вы намерены сделать бизнес. Так как я не знаю (и, возможно, вы еще не знаете), что это будет за отрасль, я не могу порекомендовать вам книги по вашему предмету.
Но есть две очень полезные книги, в которых рассказано, как стать лучшим в том, чем вы занимаетесь (чем бы вы не занимались):
ЧАВО
В заключение я также отвечу на несколько часто задаваемых вопросов, которые возникают у молодых людей, стремящихся стать предпринимателями.
На кого учиться, чтобы стать предпринимателем?
Стать предпринимателем можно с абсолютно любым образованием, хоть с медицинским, хоть с филологическим. Так что просто выбирайте, что вам больше нравится. В чем, как вам кажется, вы сможете хорошо разобраться и стать экспертом. В идеале, в числе доступных вам для обучения курсов должен быть маркетинг, но сегодня доступно столько хороших книг, столько видео и курсов, что любые недостающие к вашей программе знания вы наберете без труда.
Впрочем, некоторые высшие учебные заведения учат на “предпринимателя”, например, уже упомянутый мною Университет Синергия.
Стоит ли учиться на предпринимателя сразу после 11 класса?
Те источники информации, которые я рекомендую в этой статье, имеет смысл читать и смотреть и после 11 класса, и даже в 10 – 11 классе. Вредно не будет. Если вы еще не закончили школу, начните с книг Ричарда Брэнсона и Тины Силиг.
 instagram.com/p/BW5NO5CllbS/» data-instgrm-version=»8″>
instagram.com/p/BW5NO5CllbS/» data-instgrm-version=»8″>Что касается самой специальности “предприниматель”, если вы хотите учиться именно ей, пожалуй, ее лучше взять как магистерскую специальность после бакалавриата по маркетингу.
На кого нужно учиться, чтобы иметь больше всего шансов преуспеть в любом бизнесе?
Маркетолог – это та профессия, которая является “ключом” к любому бизнесу. Если вы не хотите жестко специализировать и быть программистом или инженером, но хотите иметь возможность работать в любом бизнесе, учитесь на маркетолога. Маркетинг и продажи – это 50% успеха абсолютно в любой отрасли.
Так что учитесь на маркетолога, поработайте во время учебы продавцом (можно продавать рекламу в журнале или на радио или продавать бытовую технику, не важно), и вы получите прекрасный фундамент для успеха в бизнесе.
Где учиться на предпринимателя или на маркетолога?
Молодым людям я рекомендую Университет Синергия или Высшую школу экономики. Есть и другие сильные Вузы, но если говорить об университетах для будущих предпринимателей, моя рекомендация такая.
Есть и другие сильные Вузы, но если говорить об университетах для будущих предпринимателей, моя рекомендация такая.
Для состоявшихся профессионалов с опытом работы лучшим российским вариантом будет бизнес-школа Сколково.
Существует ли официальная профессия “предприниматель”?
В России есть официальный классификатор профессий и должностей (ОКПДТР), в нем собраны все должности и профессии, которые официально существуют в стране, включая профессию “президент”. Профессия “предприниматель” в нем пока отсутствует. Официально предприниматель – это статус (“индивидуальный предприниматель”), а не профессия.
Неофициально предприниматель – это все же профессия, в любых документах, в частности, в анкете при заявлении на визу можно указывать “предприниматель” в качестве профессии.
Сколько учиться, чтобы достичь высот в бизнесе?
Есть универсальное правило “10 000 часов”, чтобы достичь выдающихся высот в какой-либо сфере деятельности или отрасли знаний, нужно заниматься ей около 10 000 часов.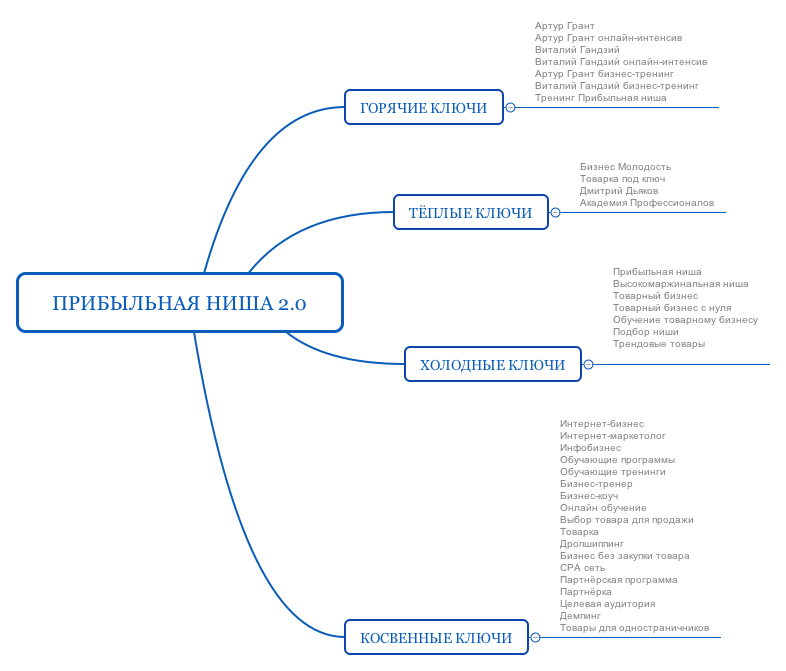 В случае с предпринимателем речь идет, конечно, не только об обучении, но и о практике.
В случае с предпринимателем речь идет, конечно, не только об обучении, но и о практике.
О статье
Название
Где учиться, чтобы стать предпринимателем » На кого учиться ✔️ Университеты ✔️ Тренинги
Анонс
Где учиться ► Чтобы стать предпринимателем ✅ Университеты и институты ✅ Курсы и тренинги ✅ Книги о бизнесе ✅ Что смотреть на youtube ✅ На кого подписаться ✅ Как развивать мышление предпринимателя
Автор
Артем Аношкин
Сайт
Инвестиции в себя — InvestmentRussia.Ru
Ниша костного мозга для гемопоэтических стволовых клеток
Природа. Авторская рукопись; Доступен в PMC 2015 JUL 24.
Опубликовано в окончательной редактированной форме AS:
PMCID: PMC4514480
NIHMSID: NIHMS708815
Sean J. Morrison
1 Говард Хьюзский медицинский институт, детский научно-исследовательский институт Педиатрия, Юго-западный медицинский центр Техасского университета, Даллас, Техас, 75390, США
Дэвид Т.
Скадден2 Центр регенеративной медицины Массачусетской больницы общего профиля, Гарвардский институт стволовых клеток и отделение стволовых клеток и регенеративной биологии, Гарвардский университет, Кембридж, Массачусетс, 02138, США
1 Медицинский институт Говарда Хьюза, Детский научно-исследовательский институт, кафедра педиатрии, Юго-западный медицинский центр Техасского университета, Даллас, Техас, 75390, США
2 Центр регенеративной медицины , Больница общего профиля Массачусетса, Гарвардский институт стволовых клеток и отделение стволовых клеток и регенеративной биологии , Гарвардский университет, Кембридж, Массачусетс 02138, США
Окончательная отредактированная версия этой статьи доступна на сайте Nature См. другие статьи PMC, в которых цитируется опубликованная статья.Предисловие
Ниши представляют собой локальные тканевые микроокружения, которые поддерживают и регулируют стволовые клетки. Гематопоэз обеспечивает парадигму для понимания стволовых клеток млекопитающих и их ниш, однако ниша гемопоэтических стволовых клеток (HSC) остается не полностью определенной и подвергается воздействию конкурирующих моделей. Здесь мы рассматриваем прогресс в выяснении местоположения и клеточных компонентов ниши HSC в костном мозге. Ниша является периваскулярной, частично образована мезенхимальными стромальными и эндотелиальными клетками и часто, но не всегда, расположена вблизи трабекулярной кости.Нерешенные вопросы касаются клеточной сложности ниши, роли эндоста и функциональной гетерогенности периваскулярного микроокружения.
Здесь мы рассматриваем прогресс в выяснении местоположения и клеточных компонентов ниши HSC в костном мозге. Ниша является периваскулярной, частично образована мезенхимальными стромальными и эндотелиальными клетками и часто, но не всегда, расположена вблизи трабекулярной кости.Нерешенные вопросы касаются клеточной сложности ниши, роли эндоста и функциональной гетерогенности периваскулярного микроокружения.
Введение
Ниши HSC присутствуют в различных тканях на протяжении всего развития, начиная с области аорты-гонад-мезонефроса (AGM) и желточного мешка, затем следуют плацента, печень плода, селезенка и костный мозг 1 . В постнатальном периоде костный мозг является первичным местом поддержания ГСК и кроветворения, но в ответ на гемопоэтический стресс ниша может перемещаться в экстрамедуллярные участки.Определение компонентов ниш и того, как они работают совместно для регуляции гемопоэза, дает возможность улучшить регенерацию после травмы или трансплантации HSC и понять, как нарушенная функция ниши может способствовать заболеванию. В этом обзоре мы сосредоточимся на природе ниши HSC в костном мозге, потому что это было предметом большинства недавних исследований и споров.
В этом обзоре мы сосредоточимся на природе ниши HSC в костном мозге, потому что это было предметом большинства недавних исследований и споров.
Исторический контекст
Вслед за Дарвином большое внимание уделялось определению иерархических эволюционных взаимоотношений между организмами.Морфологическое сходство использовалось для построения родовых деревьев, которые соединяли сложные многоклеточные организмы с оригинальной одноклеточной «стволовой клеткой» 2 . Были сформулированы родственные связи, и Эрнст Геккель предположил, что клеточная организация в развивающемся организме представляет собой повторение событий эволюции вида, при этом клетки происходят из «стволовой клетки», эквивалентной 3 . Тридцать лет спустя Артур Паппенгейм предложил менее грандиозную и более точную формулировку, основанную на улучшенной способности визуализировать клеточную морфологию: клетки крови связаны друг с другом, а зрелые типы клеток происходят от одного типа клеток в «едином представлении о гемопоэз» 4 . При этом он сформулировал гипотезу тканевых стволовых клеток. На экспериментальное определение этой концепции ушло примерно полвека благодаря вдохновляющей работе Тилля и МакКаллоха, которые показали, что отдельные клетки действительно могут давать многолинейных потомков, сохраняя мультипотентность материнской клетки 5–7 . Они обосновали идею стволовой клетки и дали нам методы для определения основных свойств этих клеток, самообновления и дифференцировки.
При этом он сформулировал гипотезу тканевых стволовых клеток. На экспериментальное определение этой концепции ушло примерно полвека благодаря вдохновляющей работе Тилля и МакКаллоха, которые показали, что отдельные клетки действительно могут давать многолинейных потомков, сохраняя мультипотентность материнской клетки 5–7 . Они обосновали идею стволовой клетки и дали нам методы для определения основных свойств этих клеток, самообновления и дифференцировки.
Till и McCulloch основывали большую часть своей работы на анализе колониеобразующих клеток селезенки in vivo (CFU-S), который, как теперь известно, измеряет главным образом мультипотентные клетки-предшественники, а не долгосрочные самообновляющиеся гемопоэтические стволовые клетки (HSCs) 8,9 .Неточный характер этого анализа способствовал формулировке Рэем Шофилдом гипотезы ниши в 1978 году. Признавая, что предполагаемые стволовые клетки КОЕ-С менее устойчивы, чем клетки костного мозга, при восстановлении кроветворения у облученных животных, он предположил, что специализированный костный мозг ниша сохранила восстанавливающую способность стволовых клеток 10 . Его коллеги из Манчестерского университета одновременно стремились определить, что делает костный мозг питательной средой для ГСК, и Майкл Декстер показал, что в основном мезенхимальные «стромальные» клеточные культуры могут поддерживать примитивные гемопоэтические клетки ex vivo 11 .Кроме того, Brian Lord постепенно рассверливал длинные полости костного мозга и показал, что примитивные клетки имели тенденцию локализоваться в направлении эндостальных краев, что привело к гипотезе о том, что кость может регулировать гемопоэз ( 12 ).
Его коллеги из Манчестерского университета одновременно стремились определить, что делает костный мозг питательной средой для ГСК, и Майкл Декстер показал, что в основном мезенхимальные «стромальные» клеточные культуры могут поддерживать примитивные гемопоэтические клетки ex vivo 11 .Кроме того, Brian Lord постепенно рассверливал длинные полости костного мозга и показал, что примитивные клетки имели тенденцию локализоваться в направлении эндостальных краев, что привело к гипотезе о том, что кость может регулировать гемопоэз ( 12 ).
Кроветворные стволовые клетки (ГСК) во взрослом возрасте находятся преимущественно в костном мозге. Костный мозг представляет собой сложный орган, содержащий множество различных типов гемопоэтических и негематопоэтических клеток. Костный мозг окружен оболочкой из васкуляризированной и иннервированной кости.а. Мельчайшие выступы кости (трабекулы) обнаруживаются по всему метафизу, так что многие клетки в этой области располагаются близко к поверхности кости. б. Поверхность раздела кости и костного мозга известна как эндост, который покрыт клетками, выстилающими кость, которые включают костеобразующие остеобласты и остеокласты, резорбирующие кость. Артерии переносят кислород, питательные вещества и факторы роста в костный мозг, прежде чем попасть в синусоиды, которые сливаются в центральный синус, образуя венозное кровообращение. Синусоиды представляют собой специализированные венулы, образующие ретикулярную сеть фенестрированных сосудов, которые позволяют клеткам входить и выходить из кровотока.Рядом с эндостом особенно много артериол, а также синусоидов. в. Трехмерная реконструированная микрофотография костного мозга, обращенная к эндостальной поверхности (синий цвет) с глубины 50 мкм ниже поверхности, на которой видна богатая сеть сосудов (красный цвет) (любезно предоставлено Charles Lin, Joel Spencer и Juwell Wu). Мелкие артериолярные сосуды (белые стрелки) становятся более крупными синусоидальными сосудами. Поле зрения составляет 350 мкм × 350 мкм.
б. Поверхность раздела кости и костного мозга известна как эндост, который покрыт клетками, выстилающими кость, которые включают костеобразующие остеобласты и остеокласты, резорбирующие кость. Артерии переносят кислород, питательные вещества и факторы роста в костный мозг, прежде чем попасть в синусоиды, которые сливаются в центральный синус, образуя венозное кровообращение. Синусоиды представляют собой специализированные венулы, образующие ретикулярную сеть фенестрированных сосудов, которые позволяют клеткам входить и выходить из кровотока.Рядом с эндостом особенно много артериол, а также синусоидов. в. Трехмерная реконструированная микрофотография костного мозга, обращенная к эндостальной поверхности (синий цвет) с глубины 50 мкм ниже поверхности, на которой видна богатая сеть сосудов (красный цвет) (любезно предоставлено Charles Lin, Joel Spencer и Juwell Wu). Мелкие артериолярные сосуды (белые стрелки) становятся более крупными синусоидальными сосудами. Поле зрения составляет 350 мкм × 350 мкм. д. Вид в поперечном сечении кровеносных сосудов, которые проходят вдоль эндостальной поверхности (ev) и переходят (белая стрелка) в синусоиды (s), которые затем направляются к центральному синусу (из ref 31 ).е. Костный мозг представляет собой клеточный комплекс с CD150 + CD48 — CD41 — ГСК Lineage — (стрелка), находящимися в тесном контакте не только с сосудистыми и периваскулярными клетками (*, просветы синусоидов), но и с мегакариоцитами (большие желтые клетки) и другие гемопоэтические клетки (изображение из ссылки 124 ).
д. Вид в поперечном сечении кровеносных сосудов, которые проходят вдоль эндостальной поверхности (ev) и переходят (белая стрелка) в синусоиды (s), которые затем направляются к центральному синусу (из ref 31 ).е. Костный мозг представляет собой клеточный комплекс с CD150 + CD48 — CD41 — ГСК Lineage — (стрелка), находящимися в тесном контакте не только с сосудистыми и периваскулярными клетками (*, просветы синусоидов), но и с мегакариоцитами (большие желтые клетки) и другие гемопоэтические клетки (изображение из ссылки 124 ).
За этими ранними исследованиями последовали доказательства in vitro , что остеобласты , дифференцированные в культуре из стромальных клеток костного мозга человека , могут продуцировать гемопоэтические цитокины и поддерживать примитивные гемопоэтические клетки в культуре 13 .Это подтолкнуло к идее, что костные клетки могут создать нишу HSC, но было важно перейти к сконструированным линиям мышей, чтобы проверить гипотезу in vivo. Затем последовали два исследования, включая модель на мышах, в которой промотор, ограниченный по активности остеобластическими клетками, использовался для управления экспрессией конститутивно активного рецептора паратиреоидного гормона 14 . Аналогичным образом, лаборатория Linheng Li использовала промотор, поскольку было показано, что он ограничен в строме костного мозга примитивными и зрелыми клетками остеолинии 15 для удаления гена BMPr1a 16 .В обеих моделях количество эндостальных остеобластов и количество примитивных гемопоэтических клеток (рассчитанных как стволовые клетки с учетом используемых в то время показателей) увеличились. Эти данные предоставили первое свидетельство того, что специфические гетерологичные клетки регулируют стволовые клетки млекопитающих in vivo, хотя оставалось неясным, была ли регуляция прямой или косвенной. Это продемонстрировало, что ниша была экспериментально управляемой, что побудило к серии исследований, которые с тех пор улучшили наше понимание сложности микроокружения костного мозга.
Затем последовали два исследования, включая модель на мышах, в которой промотор, ограниченный по активности остеобластическими клетками, использовался для управления экспрессией конститутивно активного рецептора паратиреоидного гормона 14 . Аналогичным образом, лаборатория Linheng Li использовала промотор, поскольку было показано, что он ограничен в строме костного мозга примитивными и зрелыми клетками остеолинии 15 для удаления гена BMPr1a 16 .В обеих моделях количество эндостальных остеобластов и количество примитивных гемопоэтических клеток (рассчитанных как стволовые клетки с учетом используемых в то время показателей) увеличились. Эти данные предоставили первое свидетельство того, что специфические гетерологичные клетки регулируют стволовые клетки млекопитающих in vivo, хотя оставалось неясным, была ли регуляция прямой или косвенной. Это продемонстрировало, что ниша была экспериментально управляемой, что побудило к серии исследований, которые с тех пор улучшили наше понимание сложности микроокружения костного мозга.
Исследования ниши в настоящее время более точно определили компоненты, которые регулируют HSCs и, в некоторой степени, другие гемопоэтические предшественники в костном мозге. Как и в любой интерактивной системе, между клетками костного мозга существуют сложные регуляторные отношения. Возмущение в одном типе клеток, которое приводит к эффекту в другом типе клеток, не обязательно требует, чтобы взаимодействие между клетками было прямым. Теперь данные свидетельствуют о том, что ранние исследования, в которых наблюдалось влияние на частоту HSC как следствие генетических манипуляций в остеобластных клетках, отражали скорее косвенные эффекты, чем существование остеобластной ниши.Действительно, экспрессия конститутивно активного рецептора паратиреоидного гормона в остеобластах 14 , вероятно, вызывает широко распространенные изменения во многих типах клеток костного мозга, в том числе в сосудистой сети. Текущие данные свидетельствуют о том, что существуют специализированные ниши для отдельных типов гемопоэтических стволовых клеток и клеток-предшественников и что каждая ниша может быть создана множеством типов клеток, которые вносят свой вклад в ниши уникальным, а также дублирующим образом -17-. В самом деле, существует гетерогенность среди самих HSCs 18-20 , что повышает вероятность наличия клеточно различных ниш для различных субпопуляций HSCs.В этом обзоре основное внимание будет уделено текущим данным и оставшимся без ответа вопросам.
В самом деле, существует гетерогенность среди самих HSCs 18-20 , что повышает вероятность наличия клеточно различных ниш для различных субпопуляций HSCs.В этом обзоре основное внимание будет уделено текущим данным и оставшимся без ответа вопросам.
Картирование пространства костного мозга
Ниша определяется анатомией и функцией 21 — локальная тканевая микросреда, которая непосредственно поддерживает и регулирует определенный вид стволовых клеток или клеток-предшественников 22 . Определение того, какие клетки соседствуют с HSC и регулируют поддержание HSC, было затруднено из-за сложности сохранения гистологической целостности при разрезании кости, а также из-за сложности методов иммунного окрашивания, необходимых для идентификации HSC.
Идентификация маркеров, которые надежно идентифицируют HSCs in vivo, была важным шагом в определении ниши 22 . Несмотря на возможность выделения HSC с помощью проточной цитометрии в течение десятилетий 23 , идентификация HSC в тканях оставалась проблемой, поскольку комбинация иммунофлуоресцентных маркеров, используемых для выделения HSC с помощью проточной цитометрии, была слишком сложной для микроскопии. Поэтому часто использовались маркеры низкой специфичности. Например, «HSCs» были локализованы в костном мозге с использованием удерживания метки BrdU или h3B-GFP в качестве маркера 24,25 .Хотя существует подмножество HSC, которые предпочтительно сохраняют h3B-GFP и BrdU, эти маркеры сами по себе имеют очень низкую специфичность — подавляющее большинство клеток костного мозга, которые сохраняют эти метки, не являются HSC 18,19,26 .
Поэтому часто использовались маркеры низкой специфичности. Например, «HSCs» были локализованы в костном мозге с использованием удерживания метки BrdU или h3B-GFP в качестве маркера 24,25 .Хотя существует подмножество HSC, которые предпочтительно сохраняют h3B-GFP и BrdU, эти маркеры сами по себе имеют очень низкую специфичность — подавляющее большинство клеток костного мозга, которые сохраняют эти метки, не являются HSC 18,19,26 .
Комбинируя положительное окрашивание на CD150 и отрицательное окрашивание на CD48 и CD41, HSC наконец можно было высоко очистить с помощью простого двухцветного окрашивания 27 . Все серийно трансплантируемые ГСК молодым взрослым мышам содержатся в популяции клеток костного мозга CD150 + CD48 — CD41 -/low , включая наиболее неактивные ГСК 26–29 .Это позволило локализовать ГСК в срезах кроветворных тканей с использованием проверенных маркеров высокой чистоты. Большинство клеток CD150 + CD48 — CD41 — Lineage — клеток костного мозга и селезенки локализуются рядом с синусоидными сосудами, и почти все они находятся в пределах 5 диаметров клеток синусоиды () 27,30 . Действительно, HSCs в пять раз чаще, чем другие гемопоэтические клетки, непосредственно примыкают к синусоиду 30 . ГСК распределены по всему костному мозгу, менее 20% находятся в пределах 10 мкм от эндоста 27,30–32 .Тем не менее, большинство HSC обнаруживаются в трабекулярной области костного мозга, что указывает на то, что HSC или их ниша могут прямо или косвенно регулироваться факторами, присутствующими вблизи поверхности костей.
Действительно, HSCs в пять раз чаще, чем другие гемопоэтические клетки, непосредственно примыкают к синусоиду 30 . ГСК распределены по всему костному мозгу, менее 20% находятся в пределах 10 мкм от эндоста 27,30–32 .Тем не менее, большинство HSC обнаруживаются в трабекулярной области костного мозга, что указывает на то, что HSC или их ниша могут прямо или косвенно регулироваться факторами, присутствующими вблизи поверхности костей.
а. Срезы костного мозга мышей Scf gfp/+ показывают, что нишевые клетки HSC (зеленые) включают мезенхимальные стромальные клетки и эндотелиальные клетки, которые окружают синусоиды и потенциально другие кровеносные сосуды по всему костному мозгу 64 .б – д. Большое увеличение показывает, что Scf- GFP перекрывается с эндотелиальным маркером эндоглином, но также выходит за пределы эндоглина на аблюминальной стороне синусоидов, указывая на экспрессию мезенхимальными стромальными клетками. е. Scf- GFP не экспрессируется остеопонтин+ клетками выстилки кости вокруг трабекулярной кости, но экспрессируется некоторыми близлежащими периваскулярными клетками. ф. Cxcl12- DsRed проявляет сходный паттерн экспрессии, в первую очередь периваскулярными мезенхимальными клетками и эндотелиальными клетками вокруг синусоидов по всему костному мозгу, в паттерне, который сильно перекрывается с Scf- GFP в Cxcl12 DsRed/+ ; Scf gfp/+ мышей 17 .g–j, клетки с отрицательным маркером CD150+ (g) и CD48 и Lineage (h) обычно находятся непосредственно рядом с периваскулярными клетками Scf- GFP+ (i) в костном мозге (см. j для слияния). Изображения взяты из ссылок 17,64 .
е. Scf- GFP не экспрессируется остеопонтин+ клетками выстилки кости вокруг трабекулярной кости, но экспрессируется некоторыми близлежащими периваскулярными клетками. ф. Cxcl12- DsRed проявляет сходный паттерн экспрессии, в первую очередь периваскулярными мезенхимальными клетками и эндотелиальными клетками вокруг синусоидов по всему костному мозгу, в паттерне, который сильно перекрывается с Scf- GFP в Cxcl12 DsRed/+ ; Scf gfp/+ мышей 17 .g–j, клетки с отрицательным маркером CD150+ (g) и CD48 и Lineage (h) обычно находятся непосредственно рядом с периваскулярными клетками Scf- GFP+ (i) в костном мозге (см. j для слияния). Изображения взяты из ссылок 17,64 .
Частая локализация ГСК рядом с кровеносными сосудами предполагает, что ГСК могут поддерживаться в периваскулярной нише эндотелиальными или периваскулярными клетками 27,33 . Тем не менее, HSC мобильны, регулярно входят и выходят из обращения 34 . Это повысило вероятность того, что клетки, наблюдаемые вблизи сосудов, находились в пути, возможно, задерживались при входе или выходе из кровотока в процессе миграции через сосудистые барьеры. Этот вопрос не может быть решен гистологическим анализом, который фиксирует один момент времени.
Это повысило вероятность того, что клетки, наблюдаемые вблизи сосудов, находились в пути, возможно, задерживались при входе или выходе из кровотока в процессе миграции через сосудистые барьеры. Этот вопрос не может быть решен гистологическим анализом, который фиксирует один момент времени.
Последовательная визуализация мышей с высоким разрешением оценивала трехмерное положение клеток свода черепа с течением времени 32,35 . Эти исследования показали, что примитивные гемопоэтические клетки перемещаются в специфические микродомены кровеносных сосудов костного мозга, где в изобилии присутствуют CXCL12 и E-selectin, затем остаются в этих положениях в течение недель, генерируя новые клетки, на что указывает разделение цитозольного красителя.Когда HSC визуализировали после трансплантации облученным мышам, они предпочтительно локализовались вблизи эндоста, что согласуется с тем, что эта область особенно важна для поддержания HSC 32,36 . Однако впоследствии стало известно, что облучение разрушает синусоиды в костном мозге 37 , что повышает вероятность того, что единственными кровеносными сосудами, сохранившимися после облучения, являются артериолярные сосуды вблизи эндоста. Таким образом, периэндостальная локализация ГСК в этих экспериментах могла частично отражать разрушение синусоидальных ниш облучением.В целом, данные о локализации подчеркивают возможность наличия периваскулярной ниши. Как это могло быть разрешено с историческими данными, предполагающими, что эндост и остеобласты были участниками ниши?
Таким образом, периэндостальная локализация ГСК в этих экспериментах могла частично отражать разрушение синусоидальных ниш облучением.В целом, данные о локализации подчеркивают возможность наличия периваскулярной ниши. Как это могло быть разрешено с историческими данными, предполагающими, что эндост и остеобласты были участниками ниши?
Остеобласты: более предвестник, чем хозяин
В то время как остеобластные клетки были первой клеточной популяцией, которая, как было показано, влияет на частоту гемопоэтических стволовых клеток/клеток-предшественников при возмущении in vivo 14,16 , несколько линий доказательств вызвали опасения, что эффект может быть непрямым .Во-первых, визуализирующие исследования in vivo с использованием подтвержденных маркеров или меченых стволовых клеток обнаружили несколько HSC, контактирующих с остеобластическими клетками 27,33,38,31,32 . Во-вторых, исследования, в которых истощались остеобласты при дефиците 30 бигликана или лечении дифтерийным токсином 39,40 (Scadden, неопубликовано) или увеличивались остеобласты при лечении стронцием 41 , не оказывали острого влияния на частоту ГСК.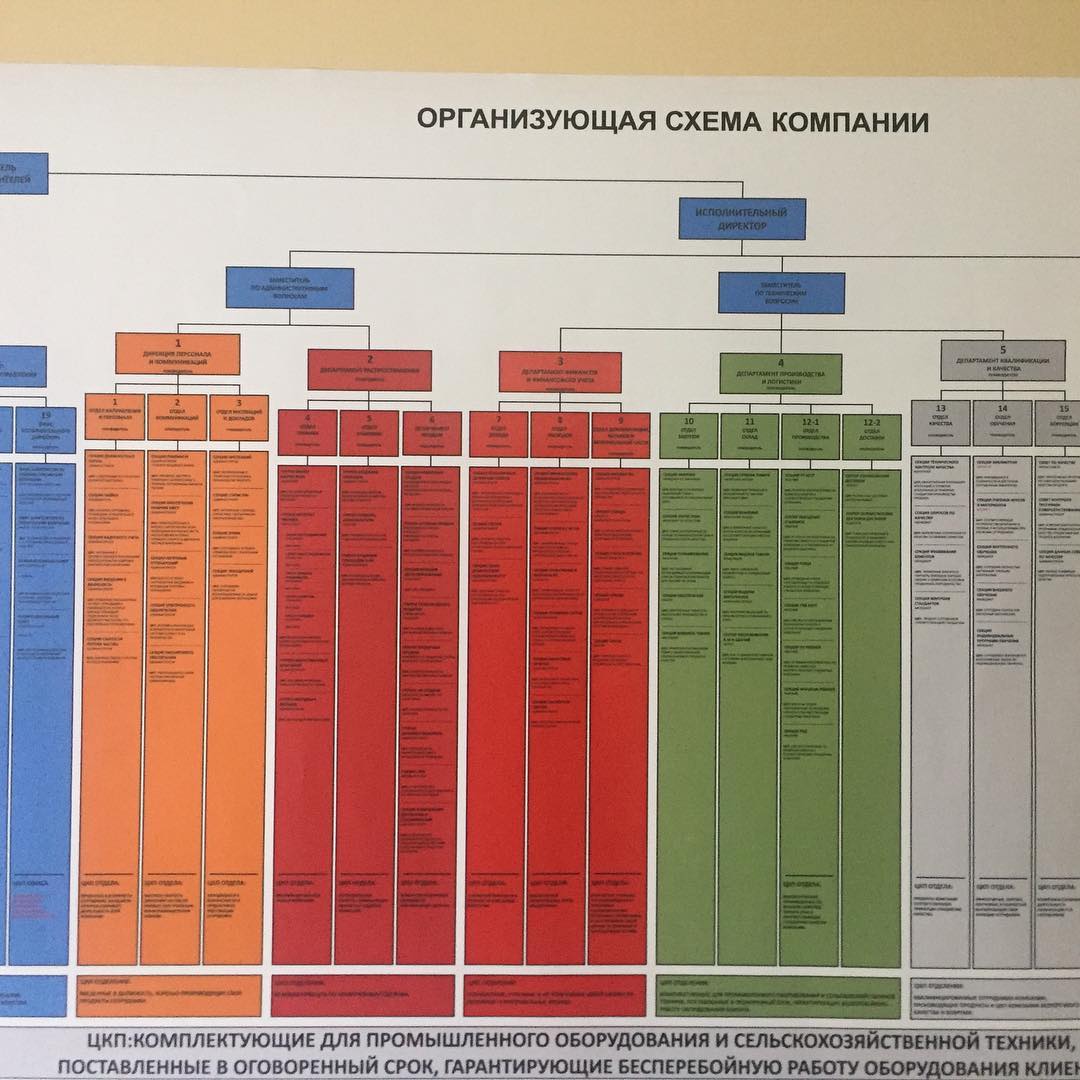 Исследования, в которых остеобласты были условно делетированы дифтерийным токсином, были особенно убедительными, поскольку они показали острое истощение лимфоидных предшественников, но не HSC 39,40 (Scadden, неопубликовано).В-третьих, генетическая модификация примитивных клеток остеолинии оказывала влияние на пролиферацию и дифференцировку HSC, но такая же модификация в зрелых остеобластах не 42 . Наконец, ключевая молекула адгезии, которая, как считается, опосредует взаимодействие остеобласт-HSC, N-cadherin, была поставлена под сомнение.
Исследования, в которых остеобласты были условно делетированы дифтерийным токсином, были особенно убедительными, поскольку они показали острое истощение лимфоидных предшественников, но не HSC 39,40 (Scadden, неопубликовано).В-третьих, генетическая модификация примитивных клеток остеолинии оказывала влияние на пролиферацию и дифференцировку HSC, но такая же модификация в зрелых остеобластах не 42 . Наконец, ключевая молекула адгезии, которая, как считается, опосредует взаимодействие остеобласт-HSC, N-cadherin, была поставлена под сомнение.
Было предложено, чтобы N-кадгерин+ HSC прикреплялись к N-кадгерин+ остеобластам посредством гомофильной адгезии 16,36 , способствуя поддержанию HSC 43–45 ; однако в этих исследованиях не проверяли, влияет ли делеция N-кадгерина на функцию HSC.Уровни окрашивания N-кадгерина в HSCs было трудно отличить от фоновой флуоресценции и зависели от антител против N-кадгерина, которые давали неспецифическое окрашивание в некоторых гемопоэтических клетках 46 . В других исследованиях не удалось обнаружить экспрессию N-кадгерина с помощью HSCs с использованием профилирования экспрессии генов множественные антитела против N-кадгерина, вестерн-блоттинг или N-кадгерин:LacZ генная ловушка мыши 18,28,30,38 .Условная делеция N-кадгерина из HSC или из клеток линии остеобластов не влияла на частоту HSC, функцию HSC или гемопоэз 38,49,50 . В совокупности эти данные подорвали представление об N-кадгерин+ «остеобластной» нише.
В других исследованиях не удалось обнаружить экспрессию N-кадгерина с помощью HSCs с использованием профилирования экспрессии генов множественные антитела против N-кадгерина, вестерн-блоттинг или N-кадгерин:LacZ генная ловушка мыши 18,28,30,38 .Условная делеция N-кадгерина из HSC или из клеток линии остеобластов не влияла на частоту HSC, функцию HSC или гемопоэз 38,49,50 . В совокупности эти данные подорвали представление об N-кадгерин+ «остеобластной» нише.
Играют ли какую-либо роль остеобласты или клетки остеолинии в регуляции ГСК? Несколько линий доказательств показывают, что эта возможность остается жизнеспособной, но не так, как предполагалось изначально. Во-первых, большее количество HSC находится в богатом трабекулярным метафизе 31,51 .Это может просто отражать совместную локализацию других компонентов костного мозга с поверхностями костей; однако, условная делеция osterix приводит к хондроцитам без дифференцировки остеобластов, увеличению кровеносных сосудов и мезенхимальных предшественников в костном мозге, но фактически устраняет гемопоэз в метафизе 52 . Эти данные доказывают, что присутствие зрелых или созревающих клеток остеолинии в областях с обильным эндостом имеет решающее значение для гемопоэза. В самом деле, мезенхимальные предшественники, способные формировать кость, достаточны для создания костных косточек, которые инвестируются сосудистой сетью хозяина и HSCs 53,54 .Это указывает на то, что кость или костеобразующие предшественники могут способствовать образованию или поддержанию ниш HSC (например, рекрутируя сосудистую сеть в костный мозг), даже если они напрямую не способствуют поддержанию HSC.
Эти данные доказывают, что присутствие зрелых или созревающих клеток остеолинии в областях с обильным эндостом имеет решающее значение для гемопоэза. В самом деле, мезенхимальные предшественники, способные формировать кость, достаточны для создания костных косточек, которые инвестируются сосудистой сетью хозяина и HSCs 53,54 .Это указывает на то, что кость или костеобразующие предшественники могут способствовать образованию или поддержанию ниш HSC (например, рекрутируя сосудистую сеть в костный мозг), даже если они напрямую не способствуют поддержанию HSC.
Трансплантированные гемопоэтические стволовые клетки/клетки-предшественники предпочтительно локализуются в кровеносных сосудах в эндостальных областях даже без предварительной цитотоксической обработки 55 . Внутри эндостальной области трансплантированные HSCs располагаются ближе к эндостальной поверхности, чем клетки-предшественники -32-.Это может снова отражать косвенные эффекты костеобразующих клеток остеолинии, т. к. костный обмен приводит к высоким локальным концентрациям ионного кальция, а кальций-чувствительный рецептор способствует приживлению костного мозга HSCs во время развития или после трансплантации 56 . Клетки остеолинии также вырабатывают цитокины и белки внеклеточного матрикса, которые могут влиять на широкий спектр типов клеток, некоторые из которых могут непосредственно регулировать функцию HSC. Примером этого служит активация рецептора паратиреоидного гормона (PTHr), которая индуцирует экспрессию множества регуляторных молекул остеобластами (таких как IL-6, RANKL и Jagged1), которые могут влиять на другие клетки костного мозга, включая сосудистую сеть 14,57 .Кроме того, остеобластная экспрессия трансгенов, кодирующих антагонисты Wnt, Dkk1 и Wif1, истощает HSCs 58,59 . Наконец, истощение экспрессирующих остеокальцин клеток (остеобластов или остеоцитов) in vivo привело к неспособности мобилизовать, по крайней мере, кратковременную репопуляцию клеток в кровь с использованием G-CSF 60,61 , несмотря на то, что остеобласты имеют небольшую экспрессию CXCL12 17, 33,62 , ключевая молекула, которая регулирует мобилизацию HSC.
к. костный обмен приводит к высоким локальным концентрациям ионного кальция, а кальций-чувствительный рецептор способствует приживлению костного мозга HSCs во время развития или после трансплантации 56 . Клетки остеолинии также вырабатывают цитокины и белки внеклеточного матрикса, которые могут влиять на широкий спектр типов клеток, некоторые из которых могут непосредственно регулировать функцию HSC. Примером этого служит активация рецептора паратиреоидного гормона (PTHr), которая индуцирует экспрессию множества регуляторных молекул остеобластами (таких как IL-6, RANKL и Jagged1), которые могут влиять на другие клетки костного мозга, включая сосудистую сеть 14,57 .Кроме того, остеобластная экспрессия трансгенов, кодирующих антагонисты Wnt, Dkk1 и Wif1, истощает HSCs 58,59 . Наконец, истощение экспрессирующих остеокальцин клеток (остеобластов или остеоцитов) in vivo привело к неспособности мобилизовать, по крайней мере, кратковременную репопуляцию клеток в кровь с использованием G-CSF 60,61 , несмотря на то, что остеобласты имеют небольшую экспрессию CXCL12 17, 33,62 , ключевая молекула, которая регулирует мобилизацию HSC. В совокупности эти данные указывают на то, что эндостальная область важна для гемопоэза, но зрелые клетки остеолинии, вероятно, играют косвенную роль в модуляции HSCs.Скорее эти клетки, по-видимому, более важны для непосредственной регуляции ограниченных предшественников, тема обсуждается ниже.
В совокупности эти данные указывают на то, что эндостальная область важна для гемопоэза, но зрелые клетки остеолинии, вероятно, играют косвенную роль в модуляции HSCs.Скорее эти клетки, по-видимому, более важны для непосредственной регуляции ограниченных предшественников, тема обсуждается ниже.
Поэтому важно переориентировать внимание на эндост как регуляторную область, а не на сами остеобласты (). Эндост имеет разнообразную группу клеток и анатомических элементов, включая большое количество артериолярных и синусоидальных кровеносных сосудов () 31,32 . Клетки включают эндотелиальные клетки, а также мезенхимальные клетки с потенциалом остеолинии.Эти мезенхимальные клетки располагаются периваскулярно, но перемещаются на эндостальную поверхность, чтобы дифференцироваться в остеобласты. Недифференцированные мезенхимальные клетки вокруг кровеносных сосудов могут способствовать поддержанию ГСК по всему костному мозгу, но мезенхимальные клетки вокруг сосудов в эндостальной области могут отличаться от клеток, удаленных от эндостальных поверхностей.
а. ГСК обнаруживаются в основном рядом с синусоидов по всему костному мозгу 27,30,31,33 , где эндотелиальные клетки и мезенхимальные стромальные клетки способствуют поддержанию ГСК путем продукции SCF 64 , CXCL12 17,33,62 и, вероятно, других факторы.Подобные клетки могут также способствовать сохранению HSC вокруг других типов кровеносных сосудов, таких как артериолы. Мезенхимальные стромальные клетки могут быть идентифицированы на основе их экспрессии LEPR- CRE 64 , PRX1- CRE 62 , CXCL12- GFP 33 , или NESTIN -GFP Transgene 63 в мыши и подобные клетки, вероятно, будут идентифицированы по экспрессии CD146 у людей 54 . Эти периваскулярные стромальные клетки, которые, вероятно, включают Cxcl12-обильные ретикулярные (CAR) клетки 33 , предназначены для формирования кости in vivo, экспрессии Mx-1-Cre и перекрытия с CD45/Ter119 — PDGFRα + Sca-1 + стромальных клеток с высоким содержанием МСК в культуре 66 . б. Вполне вероятно, что другие клетки также вносят свой вклад в эту нишу, вероятно, в том числе клетки вблизи поверхности кости в областях, богатых трабекулами. Другие типы клеток, которые регулируют ниши HSC, включают симпатические нервы 91,92 , немиелинизирующие клетки Шванна (которые также являются Nestin+) 96 , макрофаги 95 , остеокласты 97 , внеклеточный матрикс 119,1 56 . Остеобласты непосредственно не способствуют поддержанию HSC, но способствуют поддержанию и, возможно, дифференцировке определенных лимфоидных предшественников путем секреции Cxcl12 и, вероятно, др. факторов 13,17,39,40 .Таким образом, коммитированные предшественники ранних клонов находятся в эндостальной нише, которая пространственно и клеточно отличается от HSCs.
б. Вполне вероятно, что другие клетки также вносят свой вклад в эту нишу, вероятно, в том числе клетки вблизи поверхности кости в областях, богатых трабекулами. Другие типы клеток, которые регулируют ниши HSC, включают симпатические нервы 91,92 , немиелинизирующие клетки Шванна (которые также являются Nestin+) 96 , макрофаги 95 , остеокласты 97 , внеклеточный матрикс 119,1 56 . Остеобласты непосредственно не способствуют поддержанию HSC, но способствуют поддержанию и, возможно, дифференцировке определенных лимфоидных предшественников путем секреции Cxcl12 и, вероятно, др. факторов 13,17,39,40 .Таким образом, коммитированные предшественники ранних клонов находятся в эндостальной нише, которая пространственно и клеточно отличается от HSCs.
Периваскулярные регуляторы ГСК
Учитывая локализацию ГСК вблизи кровеносных сосудов, было важно определить стромальные клетки, окружающие сосуды, и проверить, способствуют ли они поддержанию ГСК. Внимание было сосредоточено на мезенхимальных клетках, окружающих кровеносные сосуды по всему костному мозгу. В то время как мезенхимальная строма, вероятно, гетерогенна, и точные отношения между клетками, экспрессирующими различные маркеры, еще предстоит определить, периваскулярные мезенхимальные клетки, экспрессирующие CD146 у человека 63 , полноразмерный рецептор лептина 64 , Prx-1- Cre 62 , Osterix-Cre 62 , и индуцируемые Mx-1- Cre 15 15 , факторы, способствующие сохранению ГСК.CXCL12-обильные «ретикулярные» (CAR) клетки, прилегающие к синусоидам, впервые показали совместную локализацию с HSCs по всему костному мозгу 33 . Удаление клеток костного мозга, экспрессирующих Cxcl12 , истощает HSC, а также серьезно снижает адипогенную и остеогенную способность клеток костного мозга 65 . Скелетные стволовые клетки человека CD146+ также локализуются рядом с синусоидами в костном мозге и синтезируют высокие уровни факторов ниши HSC, фактора стволовых клеток (SCF) и CXCL12 54 .
Внимание было сосредоточено на мезенхимальных клетках, окружающих кровеносные сосуды по всему костному мозгу. В то время как мезенхимальная строма, вероятно, гетерогенна, и точные отношения между клетками, экспрессирующими различные маркеры, еще предстоит определить, периваскулярные мезенхимальные клетки, экспрессирующие CD146 у человека 63 , полноразмерный рецептор лептина 64 , Prx-1- Cre 62 , Osterix-Cre 62 , и индуцируемые Mx-1- Cre 15 15 , факторы, способствующие сохранению ГСК.CXCL12-обильные «ретикулярные» (CAR) клетки, прилегающие к синусоидам, впервые показали совместную локализацию с HSCs по всему костному мозгу 33 . Удаление клеток костного мозга, экспрессирующих Cxcl12 , истощает HSC, а также серьезно снижает адипогенную и остеогенную способность клеток костного мозга 65 . Скелетные стволовые клетки человека CD146+ также локализуются рядом с синусоидами в костном мозге и синтезируют высокие уровни факторов ниши HSC, фактора стволовых клеток (SCF) и CXCL12 54 .
Эта возможность того, что мезенхимальные стволовые/стромальные клетки (МСК) являются частью ниши ГСК, была дополнительно подтверждена Френеттом и его коллегами, которые обнаружили, что МСК в костном мозге экспрессируют трансген Nestin -GFP и локализуются вокруг кровеносных сосудов по всей кости. костный мозг 63 . HSC обычно локализуются рядом с клетками Nestin- GFP+, а клетки Nestin- GFP+ экспрессируют высокие уровни Scf и Cxcl12 . Более того, белок активации фибробластов (FAP) экспрессируется стромальными клетками костного мозга со многими характеристиками МСК, включая Cxcl12 , Scf , экспрессию PDGFRα и Sca-1 66,67 , и удаление этих FAP+ клеток приводит к гипоцеллюлярность костного мозга, анемия и истощение остеогенных клеток 68,69 .Эти исследования предоставили убедительные доказательства того, что МСК являются одним из компонентов периваскулярной ниши для ГСК.
Эндотелиальные клетки также вносят вклад в периваскулярную нишу HSC 27 . Самым ранним функциональным доказательством, подтверждающим эту возможность, было наблюдение, что условная делеция gp130 рецептора цитокинов в эндотелиальных клетках приводит к гипоцеллюлярности костного мозга и уменьшению числа HSC 70 . Ингибирование передачи сигнала VEGFR2 у облученных мышей с помощью блокирующего антитела нарушало регенерацию синусоидальных эндотелиальных клеток и предотвращало восстановление стволовых клеток/клеток-предшественников LSK, а также колониеобразующих клеток селезенки (CFU-S) 37 .Эндотелиальные клетки могут способствовать поддержанию HSC в культуре 71 , а синусоидальные эндотелиальные клетки костного мозга способствуют долговременному восстановлению экспансии HSC в культуре 72,73 . Предполагается, что Е-селектин экспрессируется исключительно эндотелиальными клетками в костном мозге, и Дефицит Е-селектина делает HSC более неподвижными и устойчивыми к облучению 74 . Эти исследования показали, что эндотелиальные клетки являются одним из компонентов ниши HSC, но не рассматривали, регулируют ли они прямо или косвенно поддержание HSC in vivo.
Самым ранним функциональным доказательством, подтверждающим эту возможность, было наблюдение, что условная делеция gp130 рецептора цитокинов в эндотелиальных клетках приводит к гипоцеллюлярности костного мозга и уменьшению числа HSC 70 . Ингибирование передачи сигнала VEGFR2 у облученных мышей с помощью блокирующего антитела нарушало регенерацию синусоидальных эндотелиальных клеток и предотвращало восстановление стволовых клеток/клеток-предшественников LSK, а также колониеобразующих клеток селезенки (CFU-S) 37 .Эндотелиальные клетки могут способствовать поддержанию HSC в культуре 71 , а синусоидальные эндотелиальные клетки костного мозга способствуют долговременному восстановлению экспансии HSC в культуре 72,73 . Предполагается, что Е-селектин экспрессируется исключительно эндотелиальными клетками в костном мозге, и Дефицит Е-селектина делает HSC более неподвижными и устойчивыми к облучению 74 . Эти исследования показали, что эндотелиальные клетки являются одним из компонентов ниши HSC, но не рассматривали, регулируют ли они прямо или косвенно поддержание HSC in vivo.
Чтобы формально идентифицировать нишевые клетки, в исследованиях изучали, какие клеточные популяции являются ключевыми источниками факторов, способствующих поддержанию HSC in vivo. Например, SCF не является клеточно-автономным для поддержания HSC in vivo 75–79 . Дифференциальный сплайсинг и протеолитическое расщепление дают мембраносвязанные и растворимые формы SCF. HSC истощены у Sl/Sl d мутантных мышей 80 , которые экспрессируют растворимый SCF, но не мембраносвязанную форму, что указывает на то, что мембраносвязанная форма необходима для поддержания HSC 81 .Важно отметить, что мыши со смесью стромальных клеток дикого типа и Sl/Sl d обнаруживают нормальный гемопоэз только в непосредственной близости от клеток дикого типа, демонстрируя, что SCF действует локально при создании ниши 82. Поскольку HSC нуждаются в межклеточном контакте с клетками, которые синтезируют SCF, ниша может быть локализована путем идентификации ключевых источников SCF для поддержания HSC.
Анализ паттерна экспрессии SCF у нокаутированных мышей Scf gfp показал, что Scf экспрессируется периваскулярно, преимущественно вокруг синусоидов по всему костному мозгу 64 .Лептин-рецептор + (Lepr) периваскулярные стромальные клетки экспрессировали самые высокие уровни Scf , а эндотелиальные клетки экспрессировали более низкие уровни. Профилирование экспрессии генов показало, что эти периваскулярные клетки, экспрессирующие Lepr, были мезенхимальными. Экспрессия Scf- GFP не может быть обнаружена в остеобластах или гемопоэтических клетках. Условная делеция Scf из периваскулярных стромальных клеток ( Lepr- Cre) или эндотелиальных клеток ( Tie2- Cre) приводит к истощению HSC 64 .Однако делеция Scf из гемопоэтических клеток ( Vav1- Cre), остеобластных клеток ( Col2.3- Cre) и экспрессирующих нестин периваскулярных стромальных клеток ( Nestin- Cre и Nestin- CreER) не повлияло на частоту HSC 64 . Эти результаты доказывают, что существует периваскулярная ниша для ГСК, в которой эндотелиальные и мезенхимальные клетки способствуют поддержанию ГСК путем синтеза ФСК (12).
Эти результаты доказывают, что существует периваскулярная ниша для ГСК, в которой эндотелиальные и мезенхимальные клетки способствуют поддержанию ГСК путем синтеза ФСК (12).
Было высказано предположение, что эндостальная область и ее остеобластные клетки представляют собой уникальную зону для поддержания покоящихся ГСК.Однако, когда Scf был условно делетирован как из эндотелиальных клеток, так и из периваскулярных мезенхимальных клеток в Lepr-Cre; Tie2-Cre; Scf fl/ — мышей, 85% всех долгоживущих многолинейных реконструирующих клеток, включая все серийно трансплантируемые HSC и все HSC в наиболее неактивной субпопуляции, были элиминированы 20 . Следовательно, даже самые примитивные и покоящиеся ГСК поддерживаются периваскулярной нишей. Существуют ли функционально отличные периваскулярные ниши в разных областях костного мозга, например, в эндостальной области, остается открытым вопросом.
Синтезируются ли другие ключевые нишевые факторы преимущественно периваскулярными клетками? CXCL12 представляет собой хемокин, необходимый для поддержания HSC и удержания HSC в костном мозге 33,83–86 . Глобальная делеция Cxcl12, или гена, кодирующего его рецептор, Cxcr4 , истощает ГСК костного мозга 33,83,87 . CXCL12 в основном экспрессируется периваскулярными мезенхимальными стромальными клетками (клетки CAR, Nestin- GFP, Lepr- Cre или Prx-1- Cre, экспрессирующие клетки), при этом уровни экспрессии в 100 раз ниже в эндотелиальных клетках и 1000 -кратное снижение уровня в остеобластах 17,33,62,88,89 .Условная делеция Cxcl12 из периваскулярных мезенхимальных клеток с использованием Prx1- Cre и Lepr- Cre истощенных и мобилизованных HSC соответственно 17,62 . HSC были истощены, но не мобилизованы, когда Cxcl12 был условно делетирован из эндотелиальных клеток ( Tie2- Cre) 17,62 . Частота HSC и ретенция костного мозга не были затронуты, когда Cxcl12 был условно удален из остеобластов или их предшественников ( Col2.3- Cre и Osx- Cre), гемопоэтические клетки (Vav1-Cre) или Nestin- Cre-экспрессирующие стромальные клетки 17,62 .
Глобальная делеция Cxcl12, или гена, кодирующего его рецептор, Cxcr4 , истощает ГСК костного мозга 33,83,87 . CXCL12 в основном экспрессируется периваскулярными мезенхимальными стромальными клетками (клетки CAR, Nestin- GFP, Lepr- Cre или Prx-1- Cre, экспрессирующие клетки), при этом уровни экспрессии в 100 раз ниже в эндотелиальных клетках и 1000 -кратное снижение уровня в остеобластах 17,33,62,88,89 .Условная делеция Cxcl12 из периваскулярных мезенхимальных клеток с использованием Prx1- Cre и Lepr- Cre истощенных и мобилизованных HSC соответственно 17,62 . HSC были истощены, но не мобилизованы, когда Cxcl12 был условно делетирован из эндотелиальных клеток ( Tie2- Cre) 17,62 . Частота HSC и ретенция костного мозга не были затронуты, когда Cxcl12 был условно удален из остеобластов или их предшественников ( Col2.3- Cre и Osx- Cre), гемопоэтические клетки (Vav1-Cre) или Nestin- Cre-экспрессирующие стромальные клетки 17,62 . Эти данные подтвердили, что HSCs находятся в периваскулярной нише, в которой мезенхимальные стромальные клетки и эндотелиальные клетки синтезируют множество факторов, которые способствуют поддержанию и локализации HSC.
Эти данные подтвердили, что HSCs находятся в периваскулярной нише, в которой мезенхимальные стромальные клетки и эндотелиальные клетки синтезируют множество факторов, которые способствуют поддержанию и локализации HSC.
Хотя условное удаление Scf и Cxcl12 с Nestin- Cre и Nestin- CreER не оказало никакого влияния на частоту HSC 17,64 , Nestin ниши HSC 63 .Каждый из этих аллелей Nestin является трансгеном с различными паттернами экспрессии в костном мозге 64 . Nestin- Cre, по-видимому, не экспрессируется в костном мозге, а Nestin- CreER проявляет очень ограниченную периваскулярную экспрессию, которая не похожа на Scf- GFP, Cxcl12- DsRed, Nestin- GFP или . Nestin- Образцы экспрессии Cherry 64 . Однако экспрессия Nestin- GFP сильно перекрывается с экспрессией LepR- Cre периваскулярными клетками по всему костному мозгу 64,67,90 . Таким образом, вполне вероятно, что периваскулярные МСК Nestin- GFP+ являются компонентом ниши HSC, хотя опосредованная Nestin- Cre делеция Scf или Cxcl12 не приводит к истощению HSC. В будущем будет полезно идентифицировать другие аллели Cre, которые специфически экспрессируются в клетках Nestin- GFP+, чтобы сравнить их функцию с другими периваскулярными стромальными клетками.
Таким образом, вполне вероятно, что периваскулярные МСК Nestin- GFP+ являются компонентом ниши HSC, хотя опосредованная Nestin- Cre делеция Scf или Cxcl12 не приводит к истощению HSC. В будущем будет полезно идентифицировать другие аллели Cre, которые специфически экспрессируются в клетках Nestin- GFP+, чтобы сравнить их функцию с другими периваскулярными стромальными клетками.
Сложность периваскулярной ниши HSC
Эндотелиальные клетки и мезенхимальные стромальные клетки — не единственные типы клеток, которые регулируют периваскулярную нишу HSC ().Симпатическая нервная система регулирует экспрессию CXCL12 и задержку HSC в костном мозге 91,92 . По-видимому, это достигается симпатическими нервными волокнами, которые образуют синапсы с периваскулярными клетками вокруг подмножества кровеносных сосудов в костном мозге, обеспечивая циркадную регуляцию экспрессии CXCL12 и мобилизации HSC. Циркадные колебания клиренса старых нейтрофилов макрофагами в костном мозге также способствуют этим циркадным изменениям в экспрессии CXCL12 и циркуляции HSC 93 . В соответствии с этим, макрофаги модулируют экспрессию CXCL12 с помощью клеток Nestin- GFP+ и удержание HSC в костном мозге 94,95 . Немиелинизирующие шванновские клетки, по-видимому, регулируют нишу, регулируя активацию TGFβ и потенциально секретируя др. факторы -96-. Остеокласты или активность остеокластов в эндосте могут также влиять на поддержание HSC и удержание костного мозга 56,97,98 . Многие различные типы клеток, вероятно, прямо или косвенно регулируют периваскулярную нишу HSC.
В соответствии с этим, макрофаги модулируют экспрессию CXCL12 с помощью клеток Nestin- GFP+ и удержание HSC в костном мозге 94,95 . Немиелинизирующие шванновские клетки, по-видимому, регулируют нишу, регулируя активацию TGFβ и потенциально секретируя др. факторы -96-. Остеокласты или активность остеокластов в эндосте могут также влиять на поддержание HSC и удержание костного мозга 56,97,98 . Многие различные типы клеток, вероятно, прямо или косвенно регулируют периваскулярную нишу HSC.
Учитывая сложность типов клеток, участвующих в регуляции HSC, не существует единственной клетки-ниши. Скорее, ниша объединяет функции нескольких участников. Важно иметь в виду, что состав ниши и функция ниши могут изменяться в различных физиологических условиях или в ответ на стресс. Также важно отметить, что многие аллели рекомбиназы Cre, используемые до сих пор для изучения нишевых клеток, были активны во время развития. Хотя это было необходимо для достижения эффективной делеции гена (временно регулируемые аллели CreER имеют тенденцию давать гораздо более низкие уровни рекомбинации) и не было отмечено никаких аномалий в развитии, нельзя исключать косвенные эффекты на окружающие типы клеток и компенсаторные изменения. Хотя эндотелиальные клетки и периваскулярные мезенхимальные клетки экспрессируют SCF и CXCL12, условная делеция этих факторов из этих типов клеток может оказывать прямое и косвенное воздействие на ГСК.
Хотя эндотелиальные клетки и периваскулярные мезенхимальные клетки экспрессируют SCF и CXCL12, условная делеция этих факторов из этих типов клеток может оказывать прямое и косвенное воздействие на ГСК.
Также могут быть сигналы дальнего действия, циркулирующие через кровь, которые регулируют функцию HSC/ниши, возможно, объединяя активность стволовых клеток с общей физиологией 99 . Они могут включать гормоны, которые сигнализируют о репродуктивном или нутритивном статусе, или даже гемопоэтические цитокины. Например, тромбопоэтин необходим для поддержания HSC 100–103 .Основные места синтеза тромбопоэтина находятся в печени и почках, хотя он также синтезируется на более низких уровнях стромой костного мозга 104,105 . Эксперименты с условной делецией потребуются для определения физиологически важных источников тромбопоэтина для поддержания HSC.
В костном мозге также могут быть функционально различные периваскулярные среды в зависимости от типа сосудов. Большинство исследований периваскулярных ниш в костном мозге были сосредоточены на синусоидах, поскольку они являются наиболее многочисленными кровеносными сосудами в костном мозге, и большинство HSCs, MSCs, Scf -экспрессирующих клеток и Cxcl12 -экспрессирующих клеток находятся в непосредственной близости от их 17,27,33,54,64 .Однако другие виды кровеносных сосудов, такие как артериолы, могут играть важную роль в поддержании ГСК. Действительно, недавнее исследование показало, что NG2+, но Lepr-негативные мезенхимальные клетки, которые окружают артериолы в костном мозге, важны для поддержания покоящихся HSCs , 106, . Этот вывод был основан на наблюдении, что HSC были истощены и запущены в цикл, когда клеток NG2- CreER+ были удалены путем обработки дифтерийным токсином. Однако эти данные, по-видимому, противоречат предыдущему наблюдению о том, что покоящиеся ГСК элиминируются из костного мозга, когда Scf условно удаляется с использованием Tie2 -Cre и Lepr -Cre, которые рекомбинируют в эндотелиальных клетках и мезенхимальных клетках, которые преимущественно вокруг синусоидов по всему костному мозгу 20 .
Большинство исследований периваскулярных ниш в костном мозге были сосредоточены на синусоидах, поскольку они являются наиболее многочисленными кровеносными сосудами в костном мозге, и большинство HSCs, MSCs, Scf -экспрессирующих клеток и Cxcl12 -экспрессирующих клеток находятся в непосредственной близости от их 17,27,33,54,64 .Однако другие виды кровеносных сосудов, такие как артериолы, могут играть важную роль в поддержании ГСК. Действительно, недавнее исследование показало, что NG2+, но Lepr-негативные мезенхимальные клетки, которые окружают артериолы в костном мозге, важны для поддержания покоящихся HSCs , 106, . Этот вывод был основан на наблюдении, что HSC были истощены и запущены в цикл, когда клеток NG2- CreER+ были удалены путем обработки дифтерийным токсином. Однако эти данные, по-видимому, противоречат предыдущему наблюдению о том, что покоящиеся ГСК элиминируются из костного мозга, когда Scf условно удаляется с использованием Tie2 -Cre и Lepr -Cre, которые рекомбинируют в эндотелиальных клетках и мезенхимальных клетках, которые преимущественно вокруг синусоидов по всему костному мозгу 20 .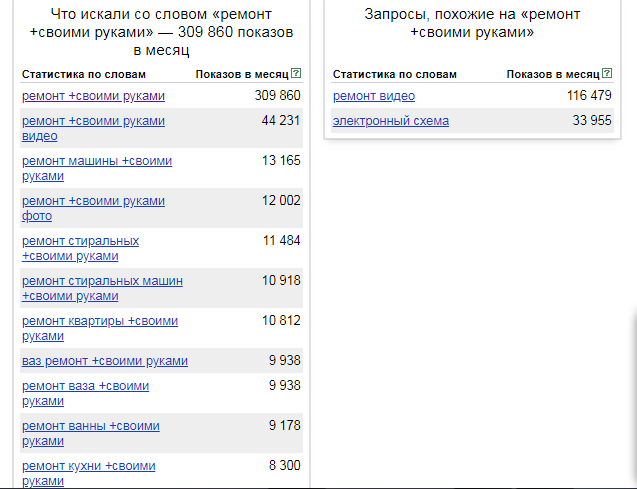 Таким образом, будет интересно определить, истощаются ли HSC, когда Scf условно делетируется с использованием NG2- CreER, или экспрессируется ли NG2- CreER клетками, отличными от периартериолярных клеток в костном мозге. Точно так же важно оценить, вносят ли периваскулярные клетки, экспрессирующие Lepr , вклад в артериолярные ниши в костном мозге. В конце концов, периваскулярные ниши, связанные как с синусоидами, так и с артериолами, могут регулировать поддержание и состояние покоя HSC в костном мозге.Анализ разнообразия в периваскулярной среде потребует аллелей Cre, которые специфически экспрессируются в пределах отдельных периваскулярных доменов, чтобы картировать их функции.
Таким образом, будет интересно определить, истощаются ли HSC, когда Scf условно делетируется с использованием NG2- CreER, или экспрессируется ли NG2- CreER клетками, отличными от периартериолярных клеток в костном мозге. Точно так же важно оценить, вносят ли периваскулярные клетки, экспрессирующие Lepr , вклад в артериолярные ниши в костном мозге. В конце концов, периваскулярные ниши, связанные как с синусоидами, так и с артериолами, могут регулировать поддержание и состояние покоя HSC в костном мозге.Анализ разнообразия в периваскулярной среде потребует аллелей Cre, которые специфически экспрессируются в пределах отдельных периваскулярных доменов, чтобы картировать их функции.
Были представлены доказательства того, что ГСК находятся в относительно гипоксических доменах костного мозга 31,107 . Это было частично основано на окрашивании пимонидазолом 31 .. Окрашенные пимонидазолом HSC часто располагаются рядом с синусоидами в костном мозге и обнаруживаются рядом с клетками, которые не окрашиваются пимонидазолом 31 .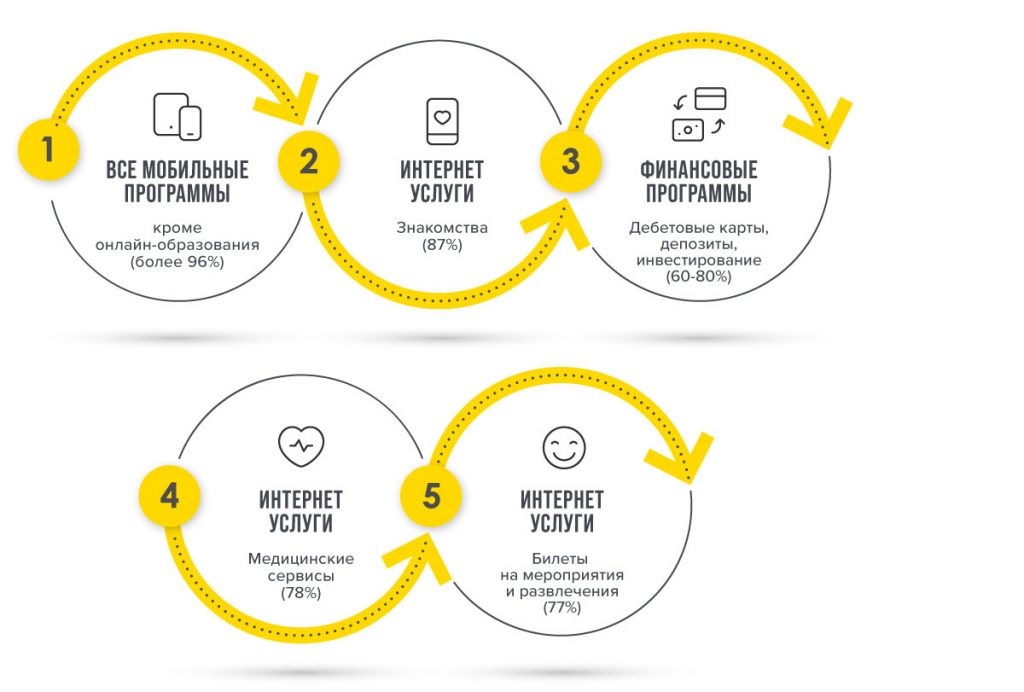 Это говорит о том, что окрашивание пимонидазолом не отражает кислород окружающей среды или что оно определяется клеточно-автономно, а не отражает гипоксическую среду. Пимонидазол реагирует на восстановление промежуточных соединений и может больше отражать метаболическое состояние клеток, чем уровень кислорода в окружающей среде.
Это говорит о том, что окрашивание пимонидазолом не отражает кислород окружающей среды или что оно определяется клеточно-автономно, а не отражает гипоксическую среду. Пимонидазол реагирует на восстановление промежуточных соединений и может больше отражать метаболическое состояние клеток, чем уровень кислорода в окружающей среде.
Зависимость поддержания HSC от HIF-1α также была интерпретирована как предположение, что HSC поддерживаются в гипоксической нише 108 . Однако ряд факторов, отличных от гипоксии, регулируют экспрессию HIF-1α.Недавнее исследование изображений с использованием нанозонда, специально отражающего окружающий кислород, показало, что напряжение кислорода было самым низким вокруг синусоидов и самым высоким вблизи эндоста (личное сообщение, Чарльз Лин). Кислород во всем пространстве костного мозга был значительно снижен по сравнению с сосудами, входящими в костный мозг, что в значительной степени утрачивалось при абляции кроветворения цитотоксическими препаратами. Поэтому вполне вероятно, что потребление кислорода во время кроветворения вызывает гипоксию костного мозга, но в эндосте не существует отчетливой гипоксической области.
Поэтому вполне вероятно, что потребление кислорода во время кроветворения вызывает гипоксию костного мозга, но в эндосте не существует отчетливой гипоксической области.
Отдельные гемопоэтические предшественники имеют разные ниши
ГСК располагаются в специализированной нише, отличной от ниш, в которых питаются другие гемопоэтические предшественники. Например, хотя остеобласты не регулируют напрямую поддержание HSC, они регулируют некоторые предшественники B-линии. Культуры, обогащенные остеобластами, поддерживают В-лимфопоэз, а удаление остеобластов у взрослых мышей приводит к острому истощению общих лимфоидных предшественников (CLP) 40,109 (Scadden, неопубликованные данные).Делеция Gs-альфа в остеобластных клетках, необходимая для передачи сигналов ПТГр, заметно истощает про- и пре-В-клетки, что может быть восстановлено с помощью IL-7 110 . Приблизительно 30% IL7R+ Lineage-клеток костного мозга, которые обогащены ранними лимфоидными предшественниками, локализуются непосредственно рядом с клетками костной выстилки в эндосте 17 . Условная делеция CXCL12, фактора, который способствует пролиферации и поддержанию предшественников В-линии 84,111 и CLP 112 , в остеобластных клетках истощает CLP и некоторые другие ранние лимфоидные предшественники из костного мозга без какого-либо влияния на HSC 17 .Следовательно, некоторые ранние лимфоидные предшественники зависят от остеобластной ниши, которая клеточно и функционально отличается от периваскулярной ниши, поддерживающей HSCs.
Условная делеция CXCL12, фактора, который способствует пролиферации и поддержанию предшественников В-линии 84,111 и CLP 112 , в остеобластных клетках истощает CLP и некоторые другие ранние лимфоидные предшественники из костного мозга без какого-либо влияния на HSC 17 .Следовательно, некоторые ранние лимфоидные предшественники зависят от остеобластной ниши, которая клеточно и функционально отличается от периваскулярной ниши, поддерживающей HSCs.
Могут существовать и другие ниши, ограниченные по происхождению. Например, макрофаги, по-видимому, имеют решающее значение для созревания эритроцитов, а истощение макрофагов снижает нормальный и злокачественный эритропоэз 113 . Другие клеточные компоненты ниши эритропоэза должны быть идентифицированы, чтобы понять взаимосвязь между этой нишей и нишами HSC и лимфоидных предшественников.
Подход условного удаления специфических нишевых факторов из клеток-кандидатов-ниш с последующим изучением последствий для поддержания стволовых клеток/клеток-предшественников in vivo дает возможность составить карту ниш для каждой стволовой клетки и ограниченной клетки-предшественника в кроветворной системе, ограниченной только точность имеющихся аллелей Cre.
Новые нишевые факторы
В отличие от стволовых клеток ряда других тканей, ГСК не могут быть устойчиво размножены в культуре.Это препятствовало возможности безопасно и эффективно трансплантировать HSC в определенных клинических контекстах, например, во время генной терапии, когда было бы полезно увеличить количество трансфицированных HSC в культуре, а затем проверить качество трансфицированных HSC перед трансплантацией. Одна возможность заключается в том, что неспособность размножать HSCs в культуре отражает существование еще не идентифицированных факторов роста, которые синтезируются нишей in vivo.
Некоторые нишевые факторы HSC были обнаружены совсем недавно.Добавление плейотрофина к культуре способствует поддержанию HSC 114 и Дефицит плейотрофина связан с истощением HSC и нарушением гемопоэтической регенерации после миелосупрессии 115 . Плейотрофин синтезируется синусоидальными эндотелиальными клетками и экспрессирующими Cxcl12 периваскулярными стромальными клетками и действует неклеточно-автономно, стимулируя функцию HSC 115 . Robo4, рецептор Slit, экспрессируемый HSCs и эндотелиальными клетками, регулирует локализацию HSC в костном мозге 116,117 .Лиганд Slit2 ограничен МСК и, возможно, другими клетками линии остеобластов. Это указывает на то, что Pleiotrophin и Robo4/Slit2 являются важными элементами периваскулярной ниши. Тенасцин-C 118 , остеопонтин 119,120 и неканонические Wnts 25 также сообщали о положительном или отрицательном влиянии на количество HSC в костном мозге и входят в число факторов, которые требуют дальнейшей характеристики с точки зрения клеточного источник или роль по отношению к нише.
Robo4, рецептор Slit, экспрессируемый HSCs и эндотелиальными клетками, регулирует локализацию HSC в костном мозге 116,117 .Лиганд Slit2 ограничен МСК и, возможно, другими клетками линии остеобластов. Это указывает на то, что Pleiotrophin и Robo4/Slit2 являются важными элементами периваскулярной ниши. Тенасцин-C 118 , остеопонтин 119,120 и неканонические Wnts 25 также сообщали о положительном или отрицательном влиянии на количество HSC в костном мозге и входят в число факторов, которые требуют дальнейшей характеристики с точки зрения клеточного источник или роль по отношению к нише.
Перспектива
Десять лет экспериментов подтвердили правильность концепции ниши и разрешили некоторые вопросы первого порядка о молекулярной и клеточной природе ниши HSC в костном мозге. Список «частей» остается неполным, но с учетом темпов текущей работы вполне вероятно, что дополнительные компоненты будут определены, а неопределенность в отношении перекрывающихся клеточных популяций разрешится в течение следующих нескольких лет. Это сделает возможным сравнение анатомически и эволюционно различных ниш HSC, которые имеют разные функции.Количество ГСК увеличивается ежедневно в печени плода, но поддерживается на почти постоянном уровне в костном мозге, по крайней мере, при отсутствии повреждений. Сравнение компонентов этих ниш может дать информацию о методах достижения расширения HSC. Точно так же сравнение гомологичных ниш среди видов, таких как долгоживущие люди и короткоживущие мыши, может дать представление о механизмах сохранения целостности кроветворения в условиях стресса или в ответ на старение. Наконец, сравнение ниш среди тканей позволит оценить, имеют ли мезенхимальные и эндотелиальные популяции в головном мозге, кишечнике и коже общие характеристики и функции с таковыми, определенными в костном мозге.Постоянно ли в различных тканях взрослых есть периваскулярные ниши для поддержания стволовых клеток? Есть ли в регенеративных тканях ниши с общими механизмами сохранения самообновления? Существуют ли общие компоненты, которые можно внедрить в ниши ex vivo?
Это сделает возможным сравнение анатомически и эволюционно различных ниш HSC, которые имеют разные функции.Количество ГСК увеличивается ежедневно в печени плода, но поддерживается на почти постоянном уровне в костном мозге, по крайней мере, при отсутствии повреждений. Сравнение компонентов этих ниш может дать информацию о методах достижения расширения HSC. Точно так же сравнение гомологичных ниш среди видов, таких как долгоживущие люди и короткоживущие мыши, может дать представление о механизмах сохранения целостности кроветворения в условиях стресса или в ответ на старение. Наконец, сравнение ниш среди тканей позволит оценить, имеют ли мезенхимальные и эндотелиальные популяции в головном мозге, кишечнике и коже общие характеристики и функции с таковыми, определенными в костном мозге.Постоянно ли в различных тканях взрослых есть периваскулярные ниши для поддержания стволовых клеток? Есть ли в регенеративных тканях ниши с общими механизмами сохранения самообновления? Существуют ли общие компоненты, которые можно внедрить в ниши ex vivo?
Теперь, когда в нашем понимании ниши костного мозга появляются подробности, можно ответить на ряд вопросов второго порядка.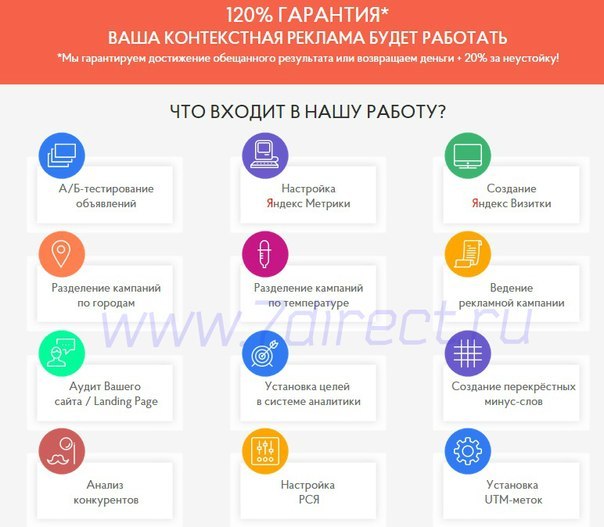 Клетки-ниши все чаще могут быть генетически помечены или модифицированы, что позволяет проводить как количественную оценку, так и молекулярные манипуляции.В сочетании с визуализацией в реальном времени с высоким разрешением и хорошо проверенными методами измерения гемопоэза становится возможным систематически выяснять, как ниша реагирует на стрессы или физиологические изменения, чтобы опосредовать изменения на уровне стволовых клеток и тканей. При стрессе из-за инфекции, миелоабляции или неоплазии, какие нишевые компоненты изменяются по количеству или функции, чтобы модифицировать гемопоэз? Существует ли иерархия нишевых компонентов, определяющих эти ответы? Может ли такая информация обеспечить прогностические алгоритмы, которые определяют конкретные вмешательства для достижения желаемых результатов?
Клетки-ниши все чаще могут быть генетически помечены или модифицированы, что позволяет проводить как количественную оценку, так и молекулярные манипуляции.В сочетании с визуализацией в реальном времени с высоким разрешением и хорошо проверенными методами измерения гемопоэза становится возможным систематически выяснять, как ниша реагирует на стрессы или физиологические изменения, чтобы опосредовать изменения на уровне стволовых клеток и тканей. При стрессе из-за инфекции, миелоабляции или неоплазии, какие нишевые компоненты изменяются по количеству или функции, чтобы модифицировать гемопоэз? Существует ли иерархия нишевых компонентов, определяющих эти ответы? Может ли такая информация обеспечить прогностические алгоритмы, которые определяют конкретные вмешательства для достижения желаемых результатов?
Другой набор вопросов касается того, каким образом ниша участвует в заболеваниях, связанных с недостаточностью стволовых клеток, таких как апластическая анемия или неоплазия.Ниша может быть враждебной по отношению к нормальным предшественникам в этих болезненных состояниях и, при неоплазии, подвергаться факультативному ответу для поддержки измененного гемопоэза 121 . Могут ли изменения в нише быть первичным, но не клеточно-автономным фактором неоплазии у людей, как было предложено на животных моделях 42,122,123 ? Потенциал для раскрытия того, как микроокружение участвует в нормальной физиологии и физиологии болезни, находится под рукой и обещает новые подходы к гематологическим нарушениям.
Могут ли изменения в нише быть первичным, но не клеточно-автономным фактором неоплазии у людей, как было предложено на животных моделях 42,122,123 ? Потенциал для раскрытия того, как микроокружение участвует в нормальной физиологии и физиологии болезни, находится под рукой и обещает новые подходы к гематологическим нарушениям.
Благодарности
SJM был поддержан Национальным институтом сердца, легких и крови (HL097760), Медицинским институтом Говарда Хьюза и кафедрой детской генетики Мэри Макдермотт Кук.DTS был поддержан Национальными институтами здравоохранения (HL044851, HL096372, EB014703) и кафедрой медицины Джеральда и Дарлин Джордан. Приносим свои извинения авторам, чьи работы не могут быть процитированы из-за нехватки места.
Ссылки
1. Миккола Х.К., Оркин С.Х. Путь разработки гемопоэтических стволовых клеток. Разработка. 2006; 133:3733–3744. doi: 10.1242/dev.02568. [PubMed] [CrossRef] [Google Scholar]2. ЭХПА Геккеля. Generale Morphologie der Organismen: allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von C. Дарвин реформирует Decendenz-Theorie. 1866. [Google Scholar]3. Геккель Э. Загадка Вселенной (Die Weltraetsel, 1895–1899) Книги Прометея; 1901. Репринтное издание 1992 г., изд. [Google Академия]5. Тилль JE, Mc CE. Прямое измерение радиационной чувствительности нормальных клеток костного мозга мыши. Радиационное разрешение 1961; 14: 213–222. [PubMed] [Google Scholar]6. Симинович Л., МакКаллок Э.А., Тилль Дж.Э. Распределение колониеобразующих клеток по колониям селезенки. J Cell Physiol. 1963; 62: 327–336. [PubMed] [Google Scholar]7.Симинович Л., Тилль Дж. Э., МакКаллок Э. А. Снижение колониеобразующей способности клеток костного мозга, подвергнутых серийной трансплантации облученным мышам. J Cell Physiol. 1964; 64: 23–31. [PubMed] [Google Scholar]8. Джонс Р.Дж. и соавт. Характеристика лимфогематопоэтических стволовых клеток мыши, лишенных колониеобразующей активности селезенки. Кровь. 1996; 88: 487–491. [PubMed] [Google Scholar]9. Спангруд Г.Дж., Брукс Д.М., Тумас Д.Б. Долговременная репопуляция облученных мышей с ограниченным количеством очищенных гемопоэтических стволовых клеток: расширение фенотипа стволовых клеток in vivo, но не функция.
Дарвин реформирует Decendenz-Theorie. 1866. [Google Scholar]3. Геккель Э. Загадка Вселенной (Die Weltraetsel, 1895–1899) Книги Прометея; 1901. Репринтное издание 1992 г., изд. [Google Академия]5. Тилль JE, Mc CE. Прямое измерение радиационной чувствительности нормальных клеток костного мозга мыши. Радиационное разрешение 1961; 14: 213–222. [PubMed] [Google Scholar]6. Симинович Л., МакКаллок Э.А., Тилль Дж.Э. Распределение колониеобразующих клеток по колониям селезенки. J Cell Physiol. 1963; 62: 327–336. [PubMed] [Google Scholar]7.Симинович Л., Тилль Дж. Э., МакКаллок Э. А. Снижение колониеобразующей способности клеток костного мозга, подвергнутых серийной трансплантации облученным мышам. J Cell Physiol. 1964; 64: 23–31. [PubMed] [Google Scholar]8. Джонс Р.Дж. и соавт. Характеристика лимфогематопоэтических стволовых клеток мыши, лишенных колониеобразующей активности селезенки. Кровь. 1996; 88: 487–491. [PubMed] [Google Scholar]9. Спангруд Г.Дж., Брукс Д.М., Тумас Д.Б. Долговременная репопуляция облученных мышей с ограниченным количеством очищенных гемопоэтических стволовых клеток: расширение фенотипа стволовых клеток in vivo, но не функция. Кровь. 1995; 85: 1006–1016. [PubMed] [Google Scholar] 10. Шофилд Р. Связь между колониеобразующей клеткой селезенки и гемопоэтической стволовой клеткой. Кровяные клетки. 1978; 4: 7–25. Артикуляция гипотезы ниши. [PubMed] [Google Scholar] 11. Декстер ТМ, Аллен ТД, Лайха ЛГ. Условия, контролирующие пролиферацию гемопоэтических стволовых клеток in vitro. J Cell Physiol. 1977; 91: 335–344. [PubMed] [Google Scholar] 12. Лорд Б.И., Теста Н.Г., Хендри Дж.Х. Относительное пространственное распределение КОЕ и КОЕ в нормальной бедренной кости мыши.Кровь. 1975; 46: 65–72. [PubMed] [Google Scholar] 13. Тайчман Р.С., Эмерсон С.Г. Остеобласты человека поддерживают гемопоэз за счет продукции гранулоцитарного колониестимулирующего фактора. J Эксперт Мед. 1994; 179: 1677–1682. [Бесплатная статья PMC] [PubMed] [Google Scholar]14. Кальви Л.М. и соавт. Остеобластические клетки регулируют нишу гемопоэтических стволовых клеток. Природа. 2003; 425:841–846. Идентификация гетерологичных клеток, влияющих на стволовые клетки/клетки-предшественники у млекопитающих, предоставляет экспериментальные доказательства гипотезы ниши.
Кровь. 1995; 85: 1006–1016. [PubMed] [Google Scholar] 10. Шофилд Р. Связь между колониеобразующей клеткой селезенки и гемопоэтической стволовой клеткой. Кровяные клетки. 1978; 4: 7–25. Артикуляция гипотезы ниши. [PubMed] [Google Scholar] 11. Декстер ТМ, Аллен ТД, Лайха ЛГ. Условия, контролирующие пролиферацию гемопоэтических стволовых клеток in vitro. J Cell Physiol. 1977; 91: 335–344. [PubMed] [Google Scholar] 12. Лорд Б.И., Теста Н.Г., Хендри Дж.Х. Относительное пространственное распределение КОЕ и КОЕ в нормальной бедренной кости мыши.Кровь. 1975; 46: 65–72. [PubMed] [Google Scholar] 13. Тайчман Р.С., Эмерсон С.Г. Остеобласты человека поддерживают гемопоэз за счет продукции гранулоцитарного колониестимулирующего фактора. J Эксперт Мед. 1994; 179: 1677–1682. [Бесплатная статья PMC] [PubMed] [Google Scholar]14. Кальви Л.М. и соавт. Остеобластические клетки регулируют нишу гемопоэтических стволовых клеток. Природа. 2003; 425:841–846. Идентификация гетерологичных клеток, влияющих на стволовые клетки/клетки-предшественники у млекопитающих, предоставляет экспериментальные доказательства гипотезы ниши. [PubMed] [Google Scholar] 15. Парк Д. и др. Эндогенные МСК костного мозга являются динамичными, ограниченными по судьбе участниками поддержания и регенерации кости. Клеточная стволовая клетка. 2012;10:259–272. doi: 10.1016/j.stem.2012.02.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]16. Чжан Дж. и др. Идентификация ниши гемопоэтических стволовых клеток и контроль размера ниши. Природа. 2003; 425:836–841. Идентификация гетерологичных клеток, влияющих на стволовые клетки/клетки-предшественники у млекопитающих, что обеспечивает экспериментальное подтверждение гипотезы Ниха.[PubMed] [Google Scholar] 17. Дин Л., Моррисон С.Дж. Кроветворные стволовые клетки и ранние лимфоидные предшественники занимают разные ниши в костном мозге. Природа. 2013; 495: 231–235. Систематический анализ экспрессирующих CXCL12 клеток в костном мозге, демонстрирующий, что стволовые клетки и ограниченные предшественники зависят от клеточно различных ниш. [Бесплатная статья PMC] [PubMed] [Google Scholar]19.
[PubMed] [Google Scholar] 15. Парк Д. и др. Эндогенные МСК костного мозга являются динамичными, ограниченными по судьбе участниками поддержания и регенерации кости. Клеточная стволовая клетка. 2012;10:259–272. doi: 10.1016/j.stem.2012.02.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]16. Чжан Дж. и др. Идентификация ниши гемопоэтических стволовых клеток и контроль размера ниши. Природа. 2003; 425:836–841. Идентификация гетерологичных клеток, влияющих на стволовые клетки/клетки-предшественники у млекопитающих, что обеспечивает экспериментальное подтверждение гипотезы Ниха.[PubMed] [Google Scholar] 17. Дин Л., Моррисон С.Дж. Кроветворные стволовые клетки и ранние лимфоидные предшественники занимают разные ниши в костном мозге. Природа. 2013; 495: 231–235. Систематический анализ экспрессирующих CXCL12 клеток в костном мозге, демонстрирующий, что стволовые клетки и ограниченные предшественники зависят от клеточно различных ниш. [Бесплатная статья PMC] [PubMed] [Google Scholar]19.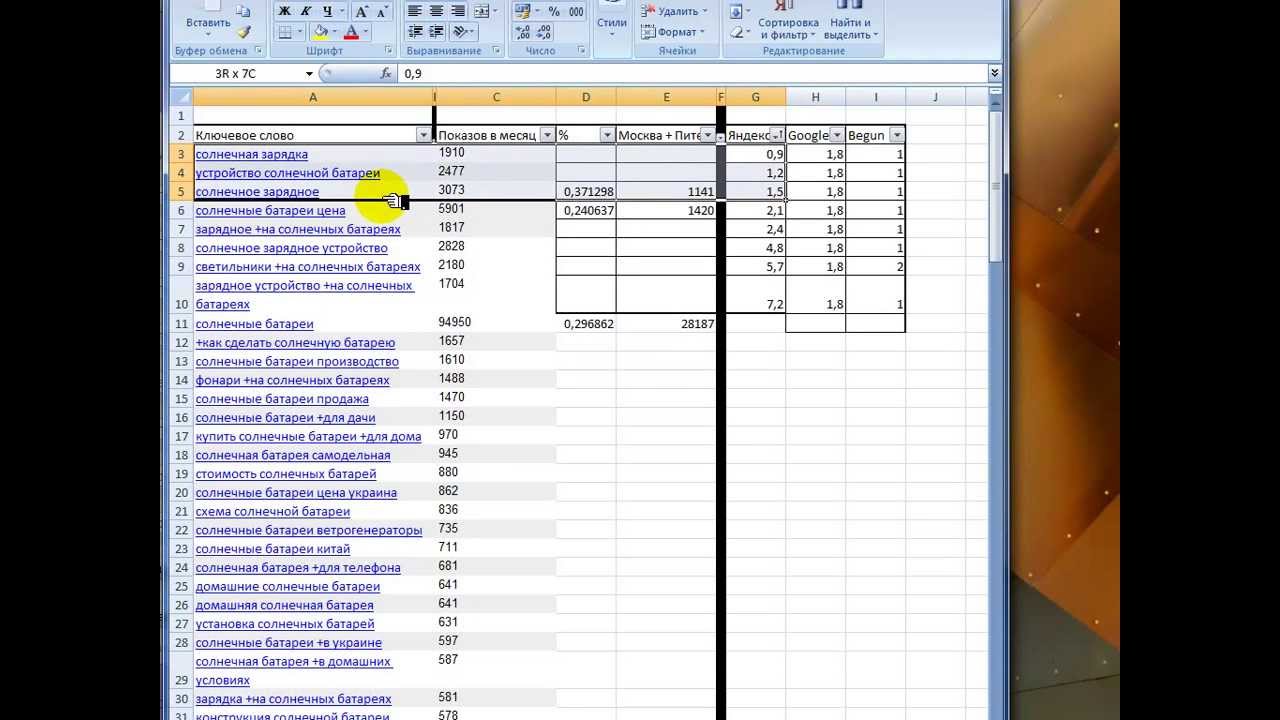 Уилсон А. и др. Гемопоэтические стволовые клетки обратимо переходят из состояния покоя в состояние самообновления в процессе гомеостаза и восстановления.Клетка. 2008; 135:1118–1129. [PubMed] [Google Scholar] 20. Огуро Х., Дин Л., Моррисон С.Дж. Маркеры семейства SLAM разрешают функционально различные субпопуляции гемопоэтических стволовых клеток и мультипотентных клеток-предшественников. Клеточная стволовая клетка. 2013; 13:102–116. doi: 10.1016/j.stem.2013.05.014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]21. Скадден ДТ. Ниша стволовых клеток как объект действия. Природа. 2006; 441:1075–1079. [PubMed] [Google Scholar] 22. Моррисон С.Дж., Спрэдлинг А.С. Стволовые клетки и ниши: механизмы, которые способствуют сохранению стволовых клеток на протяжении всей жизни.Клетка. 2008; 132: 598–611. [Бесплатная статья PMC] [PubMed] [Google Scholar]23. Spangrude GJ, Heimfeld S, Weissman IL. Очистка и характеристика гемопоэтических стволовых клеток мыши. Science (Нью-Йорк, штат Нью-Йорк, 1988; 241:58–62.
Уилсон А. и др. Гемопоэтические стволовые клетки обратимо переходят из состояния покоя в состояние самообновления в процессе гомеостаза и восстановления.Клетка. 2008; 135:1118–1129. [PubMed] [Google Scholar] 20. Огуро Х., Дин Л., Моррисон С.Дж. Маркеры семейства SLAM разрешают функционально различные субпопуляции гемопоэтических стволовых клеток и мультипотентных клеток-предшественников. Клеточная стволовая клетка. 2013; 13:102–116. doi: 10.1016/j.stem.2013.05.014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]21. Скадден ДТ. Ниша стволовых клеток как объект действия. Природа. 2006; 441:1075–1079. [PubMed] [Google Scholar] 22. Моррисон С.Дж., Спрэдлинг А.С. Стволовые клетки и ниши: механизмы, которые способствуют сохранению стволовых клеток на протяжении всей жизни.Клетка. 2008; 132: 598–611. [Бесплатная статья PMC] [PubMed] [Google Scholar]23. Spangrude GJ, Heimfeld S, Weissman IL. Очистка и характеристика гемопоэтических стволовых клеток мыши. Science (Нью-Йорк, штат Нью-Йорк, 1988; 241:58–62. [PubMed] [Google Scholar]24. Arai F, et al. Передача сигналов Tie2/ангиопоэтин-1 регулирует состояние покоя гемопоэтических стволовых клеток в нише костного мозга. Cell. 2004;118:149–161.[PubMed][Google Scholar]27.Киль М.Дж., Йилмаз О.Х., Ивашита Т., Терхорст С., Моррисон С.Дж. Рецепторы семейства SLAM различают гемопоэтические стволовые клетки и клетки-предшественники и обнаруживают эндотелиальные ниши для стволовых клеток.Клетка. 2005; 121:1109–1121. Определение иммунофенотипической сигнатуры для высокообогащенных стволовых клеток, что позволило провести гистологическое картирование HSC в костном мозге и предположить существование периваскулярной ниши. [PubMed] [Google Scholar] 28. Морита Ю., Эма Х., Накаучи Х. Гетерогенность и иерархия в самом примитивном компартменте гемопоэтических стволовых клеток. J Эксперт Мед. 2010; 207:1173–1182. doi: 10.1084/jem.200. jem.200 [pii] [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]29.Йилмаз О.Х., Киль М.Дж., Моррисон С.Дж.
[PubMed] [Google Scholar]24. Arai F, et al. Передача сигналов Tie2/ангиопоэтин-1 регулирует состояние покоя гемопоэтических стволовых клеток в нише костного мозга. Cell. 2004;118:149–161.[PubMed][Google Scholar]27.Киль М.Дж., Йилмаз О.Х., Ивашита Т., Терхорст С., Моррисон С.Дж. Рецепторы семейства SLAM различают гемопоэтические стволовые клетки и клетки-предшественники и обнаруживают эндотелиальные ниши для стволовых клеток.Клетка. 2005; 121:1109–1121. Определение иммунофенотипической сигнатуры для высокообогащенных стволовых клеток, что позволило провести гистологическое картирование HSC в костном мозге и предположить существование периваскулярной ниши. [PubMed] [Google Scholar] 28. Морита Ю., Эма Х., Накаучи Х. Гетерогенность и иерархия в самом примитивном компартменте гемопоэтических стволовых клеток. J Эксперт Мед. 2010; 207:1173–1182. doi: 10.1084/jem.200. jem.200 [pii] [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]29.Йилмаз О.Х., Киль М.Дж., Моррисон С.Дж. Маркеры семейства SLAM сохраняются среди гемопоэтических стволовых клеток старых и реконструированных мышей и заметно повышают их чистоту. Кровь. 2006; 107: 924–930. doi: 10.1182/blood-2005-05-2140. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Киль М.Дж., Радис Г.Л., Моррисон С.Дж. Отсутствие доказательств того, что гемопоэтические стволовые клетки зависят от опосредованной N-кадгерином адгезии к остеобластам для их поддержания. Клеточная стволовая клетка. 2007; 1: 204–217. [PubMed] [Google Scholar] 31.Nombela-Arrieta C, et al. Количественная визуализация локализации гемопоэтических стволовых и прогениторных клеток и гипоксического статуса в микроокружении костного мозга. Природа клеточной биологии. 2013; 15: 533–543. [Бесплатная статья PMC] [PubMed] [Google Scholar]32. Ло Селсо С. и др. Отслеживание на живых животных отдельных гемопоэтических стволовых клеток/клеток-предшественников в их нише. Природа. 2009; 457:92–96. [Бесплатная статья PMC] [PubMed] [Google Scholar]33. Сугияма Т., Кохара Х., Нода М., Нагасава Т. Поддержание пула гемопоэтических стволовых клеток с помощью передачи сигналов хемокинов CXCL12-CXCR4 в нишах стромальных клеток костного мозга.Иммунитет. 2006; 25: 977–988. doi: 10.1016/j.immuni.2006.10.016. Идентификация периваскулярной стромальной клетки, которая способствует поддержанию HSC. [PubMed] [CrossRef] [Google Scholar] 34. Райт Д.Э., Вейджерс А.Дж., Гулати А.П., Джонсон Ф.Л., Вайсман И.Л. Физиологическая миграция гемопоэтических стволовых и прогениторных клеток. Science (Нью-Йорк, штат Нью-Йорк, 2001; 294:1933–1936. [PubMed] [Google Scholar]35. Sipkins DA, et al. Визуализация in vivo специализированных эндотелиальных микродоменов костного мозга для приживления опухоли. Nature. 2005; 435: 969–973.Доказательства того, что субрегионы микроциркуляторного русла экспрессируют высокие уровни Cxcl12, где локализуются и увеличиваются количество трансплантированных гемопоэтических предшественников. [Бесплатная статья PMC] [PubMed] [Google Scholar]36.
Маркеры семейства SLAM сохраняются среди гемопоэтических стволовых клеток старых и реконструированных мышей и заметно повышают их чистоту. Кровь. 2006; 107: 924–930. doi: 10.1182/blood-2005-05-2140. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Киль М.Дж., Радис Г.Л., Моррисон С.Дж. Отсутствие доказательств того, что гемопоэтические стволовые клетки зависят от опосредованной N-кадгерином адгезии к остеобластам для их поддержания. Клеточная стволовая клетка. 2007; 1: 204–217. [PubMed] [Google Scholar] 31.Nombela-Arrieta C, et al. Количественная визуализация локализации гемопоэтических стволовых и прогениторных клеток и гипоксического статуса в микроокружении костного мозга. Природа клеточной биологии. 2013; 15: 533–543. [Бесплатная статья PMC] [PubMed] [Google Scholar]32. Ло Селсо С. и др. Отслеживание на живых животных отдельных гемопоэтических стволовых клеток/клеток-предшественников в их нише. Природа. 2009; 457:92–96. [Бесплатная статья PMC] [PubMed] [Google Scholar]33. Сугияма Т., Кохара Х., Нода М., Нагасава Т. Поддержание пула гемопоэтических стволовых клеток с помощью передачи сигналов хемокинов CXCL12-CXCR4 в нишах стромальных клеток костного мозга.Иммунитет. 2006; 25: 977–988. doi: 10.1016/j.immuni.2006.10.016. Идентификация периваскулярной стромальной клетки, которая способствует поддержанию HSC. [PubMed] [CrossRef] [Google Scholar] 34. Райт Д.Э., Вейджерс А.Дж., Гулати А.П., Джонсон Ф.Л., Вайсман И.Л. Физиологическая миграция гемопоэтических стволовых и прогениторных клеток. Science (Нью-Йорк, штат Нью-Йорк, 2001; 294:1933–1936. [PubMed] [Google Scholar]35. Sipkins DA, et al. Визуализация in vivo специализированных эндотелиальных микродоменов костного мозга для приживления опухоли. Nature. 2005; 435: 969–973.Доказательства того, что субрегионы микроциркуляторного русла экспрессируют высокие уровни Cxcl12, где локализуются и увеличиваются количество трансплантированных гемопоэтических предшественников. [Бесплатная статья PMC] [PubMed] [Google Scholar]36. Се Ю и др. Обнаружение функциональной ниши гемопоэтических стволовых клеток с использованием визуализации в реальном времени. Природа. 2008 [PubMed] [Google Scholar]37. Хупер А.Т. и соавт. Приживление и восстановление гемопоэза зависят от VEGFR2-опосредованной регенерации синусоидальных эндотелиальных клеток. Клеточная стволовая клетка.2009; 4: 263–274. doi: 10.1016/j.stem.2009.01.006. Доказательства того, что синусоидальные эндотелиальные клетки обладают специфическими свойствами и необходимы для приживления ГСК. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]38. Киль М.Дж., Акар М., Радис Г.Л., Моррисон С.Дж. Гемопоэтические стволовые клетки не зависят от N-кадгерина в регуляции их поддержания. Клеточная стволовая клетка. 2009; 4: 170–179. [Бесплатная статья PMC] [PubMed] [Google Scholar]39. Вишнич Д. и соавт. Кроветворение сильно изменено у мышей с индуцированным дефицитом остеобластов.Кровь. 2004; 103:3258–3264. [PubMed] [Google Scholar]40. Чжу Дж. и др. Остеобласты поддерживают детерминацию В-лимфоцитов и их дифференцировку из гемопоэтических стволовых клеток.
Се Ю и др. Обнаружение функциональной ниши гемопоэтических стволовых клеток с использованием визуализации в реальном времени. Природа. 2008 [PubMed] [Google Scholar]37. Хупер А.Т. и соавт. Приживление и восстановление гемопоэза зависят от VEGFR2-опосредованной регенерации синусоидальных эндотелиальных клеток. Клеточная стволовая клетка.2009; 4: 263–274. doi: 10.1016/j.stem.2009.01.006. Доказательства того, что синусоидальные эндотелиальные клетки обладают специфическими свойствами и необходимы для приживления ГСК. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]38. Киль М.Дж., Акар М., Радис Г.Л., Моррисон С.Дж. Гемопоэтические стволовые клетки не зависят от N-кадгерина в регуляции их поддержания. Клеточная стволовая клетка. 2009; 4: 170–179. [Бесплатная статья PMC] [PubMed] [Google Scholar]39. Вишнич Д. и соавт. Кроветворение сильно изменено у мышей с индуцированным дефицитом остеобластов.Кровь. 2004; 103:3258–3264. [PubMed] [Google Scholar]40. Чжу Дж. и др. Остеобласты поддерживают детерминацию В-лимфоцитов и их дифференцировку из гемопоэтических стволовых клеток. Кровь. 2007; 109:3706–3712. [PubMed] [Google Scholar]41. Лимпери С. и др. Стронций может увеличивать количество некоторых остеобластов без увеличения гемопоэтических стволовых клеток. Кровь. 2008; 111:1173–1181. doi: 10.1182/blod-2007-03-082800. [PubMed] [CrossRef] [Google Scholar]42. Raaijmakers MH, et al. Дисфункция костных предшественников вызывает миелодисплазию и вторичный лейкоз.Природа. 2010; 464: 852–857. doi: 10.1038/nature08851. Демонстрация того, что нарушение определенных мезенхимальных популяций в костном мозге может привести к патологическим гемопоэтическим исходам, включая неоплазию. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Хосокава К. и др. Нокдаун N-кадгерина подавляет длительное приживление гемопоэтических стволовых клеток. Кровь. 2010; 116: 554–563. doi: 10.1182/blood-2009-05-224857. [PubMed] [CrossRef] [Google Scholar]44. Хосокава К. и др. Адгезия на основе кадгерина является потенциальной мишенью для манипуляций с нишами для защиты гемопоэтических стволовых клеток в костном мозге взрослых.
Кровь. 2007; 109:3706–3712. [PubMed] [Google Scholar]41. Лимпери С. и др. Стронций может увеличивать количество некоторых остеобластов без увеличения гемопоэтических стволовых клеток. Кровь. 2008; 111:1173–1181. doi: 10.1182/blod-2007-03-082800. [PubMed] [CrossRef] [Google Scholar]42. Raaijmakers MH, et al. Дисфункция костных предшественников вызывает миелодисплазию и вторичный лейкоз.Природа. 2010; 464: 852–857. doi: 10.1038/nature08851. Демонстрация того, что нарушение определенных мезенхимальных популяций в костном мозге может привести к патологическим гемопоэтическим исходам, включая неоплазию. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Хосокава К. и др. Нокдаун N-кадгерина подавляет длительное приживление гемопоэтических стволовых клеток. Кровь. 2010; 116: 554–563. doi: 10.1182/blood-2009-05-224857. [PubMed] [CrossRef] [Google Scholar]44. Хосокава К. и др. Адгезия на основе кадгерина является потенциальной мишенью для манипуляций с нишами для защиты гемопоэтических стволовых клеток в костном мозге взрослых. Клеточная стволовая клетка. 2010;6:194–198. doi: 10.1016/j.stem.2009.04.013. [PubMed] [CrossRef] [Google Scholar]45. Уилсон А. и др. c-Myc контролирует баланс между самообновлением и дифференцировкой гемопоэтических стволовых клеток. Гены Дев. 2004; 18: 2747–2763. [Бесплатная статья PMC] [PubMed] [Google Scholar]46. Ли П, Зон ЛИ. Разрешение споров о N-кадгерине и гемопоэтических стволовых клетках. Клеточная стволовая клетка. 2010;6:199–202. [PubMed] [Google Scholar]47. Иванова Н.Б. и соавт. Молекулярная подпись стволовых клеток. Наука (Нью-Йорк, штат Нью-Йорк.2002; 298: 601–604. [PubMed] [Google Scholar]49. Гринбаум А.М., Револло Л.Д., Волошинек Дж.Р., Чивителли Р., Линк Д.С. N-кадгерин в клетках остеолинии не требуется для поддержания гемопоэтических стволовых клеток. Кровь. 2012 г.: 10.1182/blood-2011-09-377457. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]50. Бромберг О. и др. Остеобластический N-кадгерин не требуется для поддержки микроокружения и регуляции гемопоэтических стволовых клеток и клеток-предшественников.
Клеточная стволовая клетка. 2010;6:194–198. doi: 10.1016/j.stem.2009.04.013. [PubMed] [CrossRef] [Google Scholar]45. Уилсон А. и др. c-Myc контролирует баланс между самообновлением и дифференцировкой гемопоэтических стволовых клеток. Гены Дев. 2004; 18: 2747–2763. [Бесплатная статья PMC] [PubMed] [Google Scholar]46. Ли П, Зон ЛИ. Разрешение споров о N-кадгерине и гемопоэтических стволовых клетках. Клеточная стволовая клетка. 2010;6:199–202. [PubMed] [Google Scholar]47. Иванова Н.Б. и соавт. Молекулярная подпись стволовых клеток. Наука (Нью-Йорк, штат Нью-Йорк.2002; 298: 601–604. [PubMed] [Google Scholar]49. Гринбаум А.М., Револло Л.Д., Волошинек Дж.Р., Чивителли Р., Линк Д.С. N-кадгерин в клетках остеолинии не требуется для поддержания гемопоэтических стволовых клеток. Кровь. 2012 г.: 10.1182/blood-2011-09-377457. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]50. Бромберг О. и др. Остеобластический N-кадгерин не требуется для поддержки микроокружения и регуляции гемопоэтических стволовых клеток и клеток-предшественников. Кровь. 2012; 120:303–313. doi: 10.1182/blood-2011-09-377853.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]51. Гезгес Б. и соавт. Региональная локализация в костном мозге влияет на функциональную способность ГСК человека. Клеточная стволовая клетка. 2013;13:175–189. doi: 10.1016/j.stem.2013.06.015. [PubMed] [CrossRef] [Google Scholar]52. Чжоу С и др. Множественные функции Osterix необходимы для роста костей и гомеостаза у постнатальных мышей. Труды Национальной академии наук Соединенных Штатов Америки. 2010;107:12919–12924. дои: 10.1073/пнас.05107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]54. Саккетти Б. и др. Самообновляющиеся остеопредшественники в синусоиды костного мозга могут организовать кроветворное микроокружение. Клетка. 2007; 131:324–336. Доказательства существования мезенхимальных скелетных стволовых клеток, способных образовывать кость, секретировать нишевые факторы HSC и давать компартменты костного мозга, включающие ниши HSC. [PubMed] [Google Scholar]55. Эллис С.Л. и соавт. Связь между костью, гемопоэтическими стволовыми клетками и сосудистой сетью.Кровь. 2011; 118:1516–1524. doi: 10.1182/blood-2010-08-303800. [PubMed] [CrossRef] [Google Scholar]56. Адамс ГБ и др. Приживление стволовых клеток в эндостальной нише определяется чувствительным к кальцию рецептором. Природа. 2006; 439: 599–603. [PubMed] [Google Scholar]57. Dai JC, He P, Chen X, Greenfield EM. TNF-альфа и ПТГ используют разные механизмы для индукции экспрессии IL-6 и RANKL с заметно различающейся кинетикой. Кость. 2006; 38: 509–520. doi: 10.1016/j.bone.2005.10.007. [PubMed] [CrossRef] [Google Scholar]58.Флеминг Х.Э. и соавт. Передача сигналов Wnt в нише обеспечивает покой гемопоэтических стволовых клеток и необходима для сохранения самообновления in vivo. Клеточная стволовая клетка. 2008; 2: 274–283. [Бесплатная статья PMC] [PubMed] [Google Scholar]59. Шаниэль С. и др. Нарушение регуляции ниши костного мозга Wnt-ингибирующим фактором 1 истощает гемопоэтические стволовые клетки. Кровь. 2011;118:2420–2429. doi: 10.1182/blood-2010-09-305664. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]60. Ферраро Ф. и др. Диабет нарушает мобилизацию гемопоэтических стволовых клеток, изменяя функцию ниши.Наука трансляционная медицина. 2011;3:104ra101. doi: 10.1126/scitranslmed.3002191. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Асада Н. и др. Встроенные в матрикс остеоциты регулируют мобилизацию гемопоэтических стволовых клеток/клеток-предшественников. Клеточная стволовая клетка. 2013; 12:737–747. doi: 10.1016/j.stem.2013.05.001. [PubMed] [CrossRef] [Google Scholar]62. Гринбаум А. и др. CXCL12 у ранних мезенхимальных предшественников необходим для поддержания гемопоэтических стволовых клеток. Природа. 2013; 495: 227–230. дои: 10.1038/природа11926. Систематический анализ экспрессирующих CXCL12 клеток в костном мозге, демонстрирующий специфическую роль примитивных мезенхимальных клеток и эндотелиальных клеток в регуляции поддержания HSC. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Мендес-Феррер С. и соавт. Мезенхимальные и гемопоэтические стволовые клетки образуют уникальную нишу костного мозга. Природа. 2010; 466:829–834. doi: 10.1038/nature09262. Доказательства того, что примитивные мезенхимальные клетки, расположенные периваскулярно, регулируют поддержание ГСК.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]64. Дин Л., Сондерс Т.Л., Ениколопов Г., Моррисон С.Дж. Эндотелиальные и периваскулярные клетки поддерживают гемопоэтические стволовые клетки. Природа. 2012; 481:457–462. doi: 10.1038/nature10783. Систематический анализ экспрессии KitL в костном мозге, демонстрирующий потребность в периваскулярных клетках, экспрессирующих эндотелиальные и лептиновые рецепторы, для регуляции поддержания HSC. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Омацу Ю и др. Основные функции адипо-остеогенных клеток-предшественников как гемопоэтических стволовых клеток и ниш клеток-предшественников.Иммунитет. 2010; 33: 387–399. doi: 10.1016/j.immuni.2010.08.017. [PubMed] [CrossRef] [Google Scholar]66. Морикава С. и соавт. Проспективная идентификация, выделение и системная трансплантация мультипотентных мезенхимальных стволовых клеток в костном мозге мышей. Журнал экспериментальной медицины. 2009; 206: 2483–2496. doi: 10.1084/jem.200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Пино С. и др. PDGFR-альфа и CD51 маркируют Nestin+ сферообразующие мезенхимальные стволовые клетки человека, способные к экспансии гемопоэтических клеток-предшественников.Журнал экспериментальной медицины. 2013; 210:1351–1367. doi: 10.1084/jem.20122252. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]68. Тран Э. и др. Иммунное нацеливание на белок активации фибробластов запускает распознавание мультипотентных стромальных клеток костного мозга и кахексию. Журнал экспериментальной медицины. 2013; 210:1125–1135. doi: 10.1084/jem.20130110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]69. Робертс Э.В. и соавт. Истощение стромальных клеток, экспрессирующих белок активации фибробластов-альфа, из скелетных мышц и костного мозга приводит к кахексии и анемии.Журнал экспериментальной медицины. 2013; 210:1137–1151. doi: 10.1084/jem.20122344. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]70. Яо Л., Йокота Т., Ся Л., Кинкейд П.В., Макэвер Р.П. Дисфункция костного мозга у мышей с отсутствием цитокинового рецептора gp130 в эндотелиальных клетках. Кровь. 2005; 106:4093–4101. [Бесплатная статья PMC] [PubMed] [Google Scholar]71. Ли В., Джонсон С.А., Шелли В.К., Йодер М.К. Способность гемопоэтических стволовых клеток к репопуляции может поддерживаться in vitro некоторыми первичными эндотелиальными клетками.Экспериментальная гематология. 2004; 32:1226–1237. doi: 10.1016/j.exphem.2004.09.001. [PubMed] [CrossRef] [Google Scholar]72. Батлер Дж. М. и соавт. Эндотелиальные клетки необходимы для самообновления и репопуляции Notch-зависимых гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2010;6:251–264. doi: 10.1016/j.stem.2010.02.001. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]73. Кобаяши Х. и др. Ангиокринные факторы Akt-активированных эндотелиальных клеток уравновешивают самообновление и дифференцировку гемопоэтических стволовых клеток.Природа клеточной биологии. 2010;12:1046–1056. doi: 10.1038/ncb2108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]74. Винклер И.Г. и соавт. Сосудистая ниша Е-селектин регулирует покой гемопоэтических стволовых клеток, самообновление и химиорезистентность. Нат Мед. 2012;18:1651–1657. doi: 10.1038/nm.2969. [PubMed] [CrossRef] [Google Scholar]75. Броуди ВК. Фактор стволовых клеток и гемопоэз. Кровь. 1997; 90: 1345–1364. [PubMed] [Google Scholar]76. Чехович А., Крафт Д., Вайсман И.Л., Бхаттачарья Д. Эффективная трансплантация путем очистки ниш гемопоэтических стволовых клеток на основе антител.Science (New York, NY. 2007; 318:1296–1299. doi: 10.1126/science.1149726. [PMC free article] [PubMed] [CrossRef] [Google Scholar]78. Russell ES. Наследственные анемии мышей: a обзор для генетиков, Adv Genet, 1979, 20:357–459, [PubMed], [Google Scholar], 79. Heissig B, et al. ligand. Cell. 2002;109:625–637.[Статья PMC free] [PubMed] [Google Scholar]80. Barker JE. Гематопоэтические предшественники Sl/Sld недостаточны in situ.Экспериментальная гематология. 1994; 22: 174–177. [PubMed] [Google Scholar]81. Баркер Дж. Э. Ранняя трансплантация в нормальную микросреду предотвращает развитие дефектов гемопоэтических стволовых клеток Стилла. Экспериментальная гематология. 1997; 25: 542–547. [PubMed] [Google Scholar]82. Вольф НС. Препарирование гемопоэтического микроокружения. III. Доказательства положительного короткодействующего стимула для клеточной пролиферации. Кинет клеточной ткани. 1978; 11: 335–345. [PubMed] [Google Scholar]83. Ценг Ю.С. и соавт. Потеря Cxcl12/Sdf-1 у взрослых мышей снижает состояние покоя гемопоэтических стволовых клеток/клеток-предшественников и изменяет характер гемопоэтической регенерации после миелосупрессии.Кровь. 2011; 117:429–439. doi: 10.1182/blood-2010-01-266833. [PubMed] [CrossRef] [Google Scholar]84. Нагасава Т. и др. Дефекты В-клеточного лимфопоэза и миелопоэза костного мозга у мышей, лишенных хемокина СХС PBSF/SDF-1. Природа. 1996; 382: 635–638. [PubMed] [Google Scholar]85. Пети I и др. G-CSF индуцирует мобилизацию стволовых клеток, снижая SDF-1 костного мозга и активируя CXCR4. Нат Иммунол. 2002; 3: 687–694. [PubMed] [Google Scholar]86. Ара Т. и др. Долгоживущие гемопоэтические стволовые клетки нуждаются в факторе-1, полученном из стромальных клеток, для колонизации костного мозга во время онтогенеза.Иммунитет. 2003; 19: 257–267. [PubMed] [Google Scholar]87. Цзоу Ю.Р., Коттманн А.Х., Курода М., Таниучи И., Литтман Д.Р. Функция хемокинового рецептора CXCR4 в кроветворении и развитии мозжечка. Природа. 1998; 393: 595–599. [PubMed] [Google Scholar]88. Пономарев Т. и др. Индукция хемокинового стромального фактора-1 после повреждения ДНК улучшает функцию стволовых клеток человека. Журнал клинических исследований. 2000;106:1331–1339. doi: 10.1172/JCI10329. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]89.Дар А и др. Хемокиновый рецептор CXCR4-зависимая интернализация и ресекреция функционального хемокина SDF-1 эндотелиальными и стромальными клетками костного мозга. Нат Иммунол. 2005; 6: 1038–1046. дои: 10.1038/ni1251. [PubMed] [CrossRef] [Google Scholar]91. Катаяма Ю. и др. Сигналы от симпатической нервной системы регулируют выход гемопоэтических стволовых клеток из костного мозга. Клетка. 2006; 124:407–421. Доказательства участия нервной системы в регуляции ниши ГСК костного мозга. [PubMed] [Google Scholar]92.Мендес-Феррер С., Лукас Д., Баттиста М., Френетт П.С. Высвобождение гемопоэтических стволовых клеток регулируется циркадными колебаниями. Природа. 2008; 452:442–447. Демонстрация того, что нейронные циркадные ритмы модулируют функцию HSC. [PubMed] [Google Scholar]93. Казанова-Ацебес М. и соавт. Ритмическая модуляция гемопоэтической ниши посредством клиренса нейтрофилов. Клетка. 2013; 153:1025–1035. doi: 10.1016/j.cell.2013.04.040. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]94. Чоу А и др. Макрофаги CD169+ костного мозга способствуют удержанию гемопоэтических стволовых клеток и клеток-предшественников в нише мезенхимальных стволовых клеток.Журнал экспериментальной медицины. 2011; 208: 261–271. doi: 10.1084/jem.20101688. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]95. Винклер И.Г. и соавт. Макрофаги костного мозга поддерживают ниши гемопоэтических стволовых клеток (ГСК), и их истощение мобилизует ГСК. Кровь. 2010; 116:4815–4828. doi: 10.1182/blood-2009-11-253534. [PubMed] [CrossRef] [Google Scholar]96. Ямазаки С. и др. Немиелинизирующие шванновские клетки поддерживают гибернацию гемопоэтических стволовых клеток в нише костного мозга. Клетка. 2011; 147:1146–1158.[PubMed] [Google Scholar]97. Коллет О и др. Остеокласты разрушают эндостальные компоненты и способствуют мобилизации гемопоэтических клеток-предшественников. Нат Мед. 2006; 12: 657–664. [PubMed] [Google Scholar]98. Мансур А. и др. Остеокласты способствуют образованию ниш гемопоэтических стволовых клеток в костном мозге. Журнал экспериментальной медицины. 2012; 209: 537–549. doi: 10.1084/jem.20110994. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]99. Накада Д., Леви Б.П., Моррисон С.Дж. Интеграция физиологической регуляции с гомеостазом стволовых клеток и тканей.Нейрон. 2011;70:703–718. doi: 10.1016/j.neuron.2011.05.011. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]100. Цянь Х. и др. Критическая роль тромбопоэтина в поддержании покоя взрослых гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2007; 1: 671–684. [PubMed] [Google Scholar] 101. Йошихара Х. и др. Передача сигналов тромбопоэтина/MPL регулирует покой гемопоэтических стволовых клеток и взаимодействие с остеобластной нишей. Клеточная стволовая клетка. 2007; 1: 685–697. [PubMed] [Google Scholar] 102. Кимура С., Робертс А.В., Меткалф Д., Александр В.С.Дефицит гемопоэтических стволовых клеток у мышей, лишенных c-Mpl, рецептора тромбопоэтина. Труды Национальной академии наук США. 1998;95:1195–1200. [Бесплатная статья PMC] [PubMed] [Google Scholar]103. Каушанский К. Тромбопоэтин и гемопоэтические стволовые клетки. Кровь. 1998; 92:1–3. [PubMed] [Google Scholar] 104. Герьеро А. и др. Тромбопоэтин синтезируется стромальными клетками костного мозга. Кровь. 1997;90:3444–3455. [PubMed] [Google Scholar] 105. Сунгаран Р., Маркович Б., Чонг Б.Х. Локализация и регуляция экспрессии мРНК тромбопоэтина в почках, печени, костном мозге и селезенке человека с использованием гибридизации in situ.Кровь. 1997; 89: 101–107. [PubMed] [Google Scholar] 106. Кунисаки Ю. и др. Артериолярные ниши поддерживают гемопоэтические стволовые клетки в состоянии покоя. Природа. 2013 В прессе. Гистологическая характеристика подтипов сосудистых структур и доказательства того, что околоартериолярные мезенхимальные клетки сохраняют ГСК в состоянии покоя. [Бесплатная статья PMC] [PubMed] [Google Scholar]107. Пармар К., Маух П., Вергилио Дж. А., Сакштейн Р., Даун Д. Д. Распределение гемопоэтических стволовых клеток в костном мозге в зависимости от регионарной гипоксии. Proc Natl Acad Sci U S A.2007; 104: 5431–5436. [Бесплатная статья PMC] [PubMed] [Google Scholar]108. Такубо К. и др. Регуляция уровня HIF-1альфа необходима для гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2010;7:391–402. doi: 10.1016/j.stem.2010.06.020. [PubMed] [CrossRef] [Google Scholar] 109. Вишнич Д. и соавт. Условная абляция линии остеобластов у трансгенных мышей Col2.3deltatk. Джей Боун Шахтер Рез. 2001;16:2222–2231. [PubMed] [Google Scholar] 110. Ву Дж.И. и др. Остеобластная регуляция В-лимфопоэза опосредована Gs{альфа}-зависимыми сигнальными путями.Труды Национальной академии наук Соединенных Штатов Америки. 2008; 105:16976–16981. doi: 10.1073/pnas.0802898105. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]111. Нагасава Т., Кикутани Х., Кисимото Т. Молекулярное клонирование и структура пре-В-клеточного фактора, стимулирующего рост. Труды Национальной академии наук США. 1994; 91: 2305–2309. [Бесплатная статья PMC] [PubMed] [Google Scholar]112. Не И, Хан YC, Цзоу YR. CXCR4 необходим для покоя примитивных гемопоэтических клеток.Журнал экспериментальной медицины. 2008; 205: 777–783. doi: 10.1084/jem.20072513. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]115. Химбург Х.А. и др. Плейотрофин регулирует удержание и самообновление гемопоэтических стволовых клеток в сосудистой нише костного мозга. Cell Rep. 2012; 2:964–975. doi: 10.1016/j.celrep.2012.09.002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]116. Смит-Бердан С. и соавт. Robo4 взаимодействует с CXCR4, чтобы определить локализацию гемопоэтических стволовых клеток в нишах костного мозга.Клеточная стволовая клетка. 2011; 8:72–83. doi: 10.1016/j.stem.2010.11.030. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]117. Смит-Бердан С., Шеперс К., Ли А., Пассег Э., Форсберг Э.С. Динамическая экспрессия лиганда Robo Slit2 в популяциях клеток костного мозга. Клеточный цикл. 2012; 11: 675–682. doi: 10.4161/cc.11.4.19146. [PubMed] [CrossRef] [Google Scholar] 118. Накамура-Исидзу А. и соавт. Белок внеклеточного матрикса тенасцин-С необходим в микроокружении костного мозга, предназначенном для гемопоэтической регенерации.Кровь. 2012;119:5429–5437. doi: 10.1182/blood-2011-11-393645. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]119. Стиер С. и др. Остеопонтин является компонентом ниши гемопоэтических стволовых клеток, который негативно регулирует размер пула стволовых клеток. Журнал экспериментальной медицины. 2005; 201:1781–1791. doi: 10.1084/jem.20041992. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]120. Нильссон С.К. и др. Остеопонтин, ключевой компонент ниши гемопоэтических стволовых клеток и регулятор примитивных гемопоэтических клеток-предшественников.Кровь. 2005; 106:1232–1239. doi: 10.1182/blood-2004-11-4422. [PubMed] [CrossRef] [Google Scholar] 121. Шеперс К. и др. Миелопролиферативная неоплазия ремоделирует эндостальную нишу костного мозга в самоусиливающуюся лейкемическую нишу. Клеточная стволовая клетка. 2013; 13: 285–299. doi: 10.1016/j.stem.2013.06.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]122. Уокли Ч.Р. и др. Индуцированный микроокружением миелопролиферативный синдром, вызванный гамма-дефицитом рецептора ретиноевой кислоты. Клетка. 2007; 129:1097–1110.[Бесплатная статья PMC] [PubMed] [Google Scholar]123. Уокли Ч.Р., Ши Дж.М., Симс Н.А., Пуртон Л.Е., Оркин С.Х. Rb регулирует взаимодействие между гемопоэтическими стволовыми клетками и их микроокружением костного мозга. Клетка. 2007; 129:1081–1095. [Бесплатная статья PMC] [PubMed] [Google Scholar]124. Киль М.Дж., Моррисон С.Дж. Неопределенность в нишах, поддерживающих гемопоэтические стволовые клетки. Нат Рев Иммунол. 2008; 8: 290–301. [PubMed] [Google Scholar]
Кровь. 2012; 120:303–313. doi: 10.1182/blood-2011-09-377853.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]51. Гезгес Б. и соавт. Региональная локализация в костном мозге влияет на функциональную способность ГСК человека. Клеточная стволовая клетка. 2013;13:175–189. doi: 10.1016/j.stem.2013.06.015. [PubMed] [CrossRef] [Google Scholar]52. Чжоу С и др. Множественные функции Osterix необходимы для роста костей и гомеостаза у постнатальных мышей. Труды Национальной академии наук Соединенных Штатов Америки. 2010;107:12919–12924. дои: 10.1073/пнас.05107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]54. Саккетти Б. и др. Самообновляющиеся остеопредшественники в синусоиды костного мозга могут организовать кроветворное микроокружение. Клетка. 2007; 131:324–336. Доказательства существования мезенхимальных скелетных стволовых клеток, способных образовывать кость, секретировать нишевые факторы HSC и давать компартменты костного мозга, включающие ниши HSC. [PubMed] [Google Scholar]55. Эллис С.Л. и соавт. Связь между костью, гемопоэтическими стволовыми клетками и сосудистой сетью.Кровь. 2011; 118:1516–1524. doi: 10.1182/blood-2010-08-303800. [PubMed] [CrossRef] [Google Scholar]56. Адамс ГБ и др. Приживление стволовых клеток в эндостальной нише определяется чувствительным к кальцию рецептором. Природа. 2006; 439: 599–603. [PubMed] [Google Scholar]57. Dai JC, He P, Chen X, Greenfield EM. TNF-альфа и ПТГ используют разные механизмы для индукции экспрессии IL-6 и RANKL с заметно различающейся кинетикой. Кость. 2006; 38: 509–520. doi: 10.1016/j.bone.2005.10.007. [PubMed] [CrossRef] [Google Scholar]58.Флеминг Х.Э. и соавт. Передача сигналов Wnt в нише обеспечивает покой гемопоэтических стволовых клеток и необходима для сохранения самообновления in vivo. Клеточная стволовая клетка. 2008; 2: 274–283. [Бесплатная статья PMC] [PubMed] [Google Scholar]59. Шаниэль С. и др. Нарушение регуляции ниши костного мозга Wnt-ингибирующим фактором 1 истощает гемопоэтические стволовые клетки. Кровь. 2011;118:2420–2429. doi: 10.1182/blood-2010-09-305664. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]60. Ферраро Ф. и др. Диабет нарушает мобилизацию гемопоэтических стволовых клеток, изменяя функцию ниши.Наука трансляционная медицина. 2011;3:104ra101. doi: 10.1126/scitranslmed.3002191. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Асада Н. и др. Встроенные в матрикс остеоциты регулируют мобилизацию гемопоэтических стволовых клеток/клеток-предшественников. Клеточная стволовая клетка. 2013; 12:737–747. doi: 10.1016/j.stem.2013.05.001. [PubMed] [CrossRef] [Google Scholar]62. Гринбаум А. и др. CXCL12 у ранних мезенхимальных предшественников необходим для поддержания гемопоэтических стволовых клеток. Природа. 2013; 495: 227–230. дои: 10.1038/природа11926. Систематический анализ экспрессирующих CXCL12 клеток в костном мозге, демонстрирующий специфическую роль примитивных мезенхимальных клеток и эндотелиальных клеток в регуляции поддержания HSC. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Мендес-Феррер С. и соавт. Мезенхимальные и гемопоэтические стволовые клетки образуют уникальную нишу костного мозга. Природа. 2010; 466:829–834. doi: 10.1038/nature09262. Доказательства того, что примитивные мезенхимальные клетки, расположенные периваскулярно, регулируют поддержание ГСК.[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]64. Дин Л., Сондерс Т.Л., Ениколопов Г., Моррисон С.Дж. Эндотелиальные и периваскулярные клетки поддерживают гемопоэтические стволовые клетки. Природа. 2012; 481:457–462. doi: 10.1038/nature10783. Систематический анализ экспрессии KitL в костном мозге, демонстрирующий потребность в периваскулярных клетках, экспрессирующих эндотелиальные и лептиновые рецепторы, для регуляции поддержания HSC. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Омацу Ю и др. Основные функции адипо-остеогенных клеток-предшественников как гемопоэтических стволовых клеток и ниш клеток-предшественников.Иммунитет. 2010; 33: 387–399. doi: 10.1016/j.immuni.2010.08.017. [PubMed] [CrossRef] [Google Scholar]66. Морикава С. и соавт. Проспективная идентификация, выделение и системная трансплантация мультипотентных мезенхимальных стволовых клеток в костном мозге мышей. Журнал экспериментальной медицины. 2009; 206: 2483–2496. doi: 10.1084/jem.200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Пино С. и др. PDGFR-альфа и CD51 маркируют Nestin+ сферообразующие мезенхимальные стволовые клетки человека, способные к экспансии гемопоэтических клеток-предшественников.Журнал экспериментальной медицины. 2013; 210:1351–1367. doi: 10.1084/jem.20122252. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]68. Тран Э. и др. Иммунное нацеливание на белок активации фибробластов запускает распознавание мультипотентных стромальных клеток костного мозга и кахексию. Журнал экспериментальной медицины. 2013; 210:1125–1135. doi: 10.1084/jem.20130110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]69. Робертс Э.В. и соавт. Истощение стромальных клеток, экспрессирующих белок активации фибробластов-альфа, из скелетных мышц и костного мозга приводит к кахексии и анемии.Журнал экспериментальной медицины. 2013; 210:1137–1151. doi: 10.1084/jem.20122344. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]70. Яо Л., Йокота Т., Ся Л., Кинкейд П.В., Макэвер Р.П. Дисфункция костного мозга у мышей с отсутствием цитокинового рецептора gp130 в эндотелиальных клетках. Кровь. 2005; 106:4093–4101. [Бесплатная статья PMC] [PubMed] [Google Scholar]71. Ли В., Джонсон С.А., Шелли В.К., Йодер М.К. Способность гемопоэтических стволовых клеток к репопуляции может поддерживаться in vitro некоторыми первичными эндотелиальными клетками.Экспериментальная гематология. 2004; 32:1226–1237. doi: 10.1016/j.exphem.2004.09.001. [PubMed] [CrossRef] [Google Scholar]72. Батлер Дж. М. и соавт. Эндотелиальные клетки необходимы для самообновления и репопуляции Notch-зависимых гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2010;6:251–264. doi: 10.1016/j.stem.2010.02.001. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]73. Кобаяши Х. и др. Ангиокринные факторы Akt-активированных эндотелиальных клеток уравновешивают самообновление и дифференцировку гемопоэтических стволовых клеток.Природа клеточной биологии. 2010;12:1046–1056. doi: 10.1038/ncb2108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]74. Винклер И.Г. и соавт. Сосудистая ниша Е-селектин регулирует покой гемопоэтических стволовых клеток, самообновление и химиорезистентность. Нат Мед. 2012;18:1651–1657. doi: 10.1038/nm.2969. [PubMed] [CrossRef] [Google Scholar]75. Броуди ВК. Фактор стволовых клеток и гемопоэз. Кровь. 1997; 90: 1345–1364. [PubMed] [Google Scholar]76. Чехович А., Крафт Д., Вайсман И.Л., Бхаттачарья Д. Эффективная трансплантация путем очистки ниш гемопоэтических стволовых клеток на основе антител.Science (New York, NY. 2007; 318:1296–1299. doi: 10.1126/science.1149726. [PMC free article] [PubMed] [CrossRef] [Google Scholar]78. Russell ES. Наследственные анемии мышей: a обзор для генетиков, Adv Genet, 1979, 20:357–459, [PubMed], [Google Scholar], 79. Heissig B, et al. ligand. Cell. 2002;109:625–637.[Статья PMC free] [PubMed] [Google Scholar]80. Barker JE. Гематопоэтические предшественники Sl/Sld недостаточны in situ.Экспериментальная гематология. 1994; 22: 174–177. [PubMed] [Google Scholar]81. Баркер Дж. Э. Ранняя трансплантация в нормальную микросреду предотвращает развитие дефектов гемопоэтических стволовых клеток Стилла. Экспериментальная гематология. 1997; 25: 542–547. [PubMed] [Google Scholar]82. Вольф НС. Препарирование гемопоэтического микроокружения. III. Доказательства положительного короткодействующего стимула для клеточной пролиферации. Кинет клеточной ткани. 1978; 11: 335–345. [PubMed] [Google Scholar]83. Ценг Ю.С. и соавт. Потеря Cxcl12/Sdf-1 у взрослых мышей снижает состояние покоя гемопоэтических стволовых клеток/клеток-предшественников и изменяет характер гемопоэтической регенерации после миелосупрессии.Кровь. 2011; 117:429–439. doi: 10.1182/blood-2010-01-266833. [PubMed] [CrossRef] [Google Scholar]84. Нагасава Т. и др. Дефекты В-клеточного лимфопоэза и миелопоэза костного мозга у мышей, лишенных хемокина СХС PBSF/SDF-1. Природа. 1996; 382: 635–638. [PubMed] [Google Scholar]85. Пети I и др. G-CSF индуцирует мобилизацию стволовых клеток, снижая SDF-1 костного мозга и активируя CXCR4. Нат Иммунол. 2002; 3: 687–694. [PubMed] [Google Scholar]86. Ара Т. и др. Долгоживущие гемопоэтические стволовые клетки нуждаются в факторе-1, полученном из стромальных клеток, для колонизации костного мозга во время онтогенеза.Иммунитет. 2003; 19: 257–267. [PubMed] [Google Scholar]87. Цзоу Ю.Р., Коттманн А.Х., Курода М., Таниучи И., Литтман Д.Р. Функция хемокинового рецептора CXCR4 в кроветворении и развитии мозжечка. Природа. 1998; 393: 595–599. [PubMed] [Google Scholar]88. Пономарев Т. и др. Индукция хемокинового стромального фактора-1 после повреждения ДНК улучшает функцию стволовых клеток человека. Журнал клинических исследований. 2000;106:1331–1339. doi: 10.1172/JCI10329. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]89.Дар А и др. Хемокиновый рецептор CXCR4-зависимая интернализация и ресекреция функционального хемокина SDF-1 эндотелиальными и стромальными клетками костного мозга. Нат Иммунол. 2005; 6: 1038–1046. дои: 10.1038/ni1251. [PubMed] [CrossRef] [Google Scholar]91. Катаяма Ю. и др. Сигналы от симпатической нервной системы регулируют выход гемопоэтических стволовых клеток из костного мозга. Клетка. 2006; 124:407–421. Доказательства участия нервной системы в регуляции ниши ГСК костного мозга. [PubMed] [Google Scholar]92.Мендес-Феррер С., Лукас Д., Баттиста М., Френетт П.С. Высвобождение гемопоэтических стволовых клеток регулируется циркадными колебаниями. Природа. 2008; 452:442–447. Демонстрация того, что нейронные циркадные ритмы модулируют функцию HSC. [PubMed] [Google Scholar]93. Казанова-Ацебес М. и соавт. Ритмическая модуляция гемопоэтической ниши посредством клиренса нейтрофилов. Клетка. 2013; 153:1025–1035. doi: 10.1016/j.cell.2013.04.040. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]94. Чоу А и др. Макрофаги CD169+ костного мозга способствуют удержанию гемопоэтических стволовых клеток и клеток-предшественников в нише мезенхимальных стволовых клеток.Журнал экспериментальной медицины. 2011; 208: 261–271. doi: 10.1084/jem.20101688. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]95. Винклер И.Г. и соавт. Макрофаги костного мозга поддерживают ниши гемопоэтических стволовых клеток (ГСК), и их истощение мобилизует ГСК. Кровь. 2010; 116:4815–4828. doi: 10.1182/blood-2009-11-253534. [PubMed] [CrossRef] [Google Scholar]96. Ямазаки С. и др. Немиелинизирующие шванновские клетки поддерживают гибернацию гемопоэтических стволовых клеток в нише костного мозга. Клетка. 2011; 147:1146–1158.[PubMed] [Google Scholar]97. Коллет О и др. Остеокласты разрушают эндостальные компоненты и способствуют мобилизации гемопоэтических клеток-предшественников. Нат Мед. 2006; 12: 657–664. [PubMed] [Google Scholar]98. Мансур А. и др. Остеокласты способствуют образованию ниш гемопоэтических стволовых клеток в костном мозге. Журнал экспериментальной медицины. 2012; 209: 537–549. doi: 10.1084/jem.20110994. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]99. Накада Д., Леви Б.П., Моррисон С.Дж. Интеграция физиологической регуляции с гомеостазом стволовых клеток и тканей.Нейрон. 2011;70:703–718. doi: 10.1016/j.neuron.2011.05.011. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]100. Цянь Х. и др. Критическая роль тромбопоэтина в поддержании покоя взрослых гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2007; 1: 671–684. [PubMed] [Google Scholar] 101. Йошихара Х. и др. Передача сигналов тромбопоэтина/MPL регулирует покой гемопоэтических стволовых клеток и взаимодействие с остеобластной нишей. Клеточная стволовая клетка. 2007; 1: 685–697. [PubMed] [Google Scholar] 102. Кимура С., Робертс А.В., Меткалф Д., Александр В.С.Дефицит гемопоэтических стволовых клеток у мышей, лишенных c-Mpl, рецептора тромбопоэтина. Труды Национальной академии наук США. 1998;95:1195–1200. [Бесплатная статья PMC] [PubMed] [Google Scholar]103. Каушанский К. Тромбопоэтин и гемопоэтические стволовые клетки. Кровь. 1998; 92:1–3. [PubMed] [Google Scholar] 104. Герьеро А. и др. Тромбопоэтин синтезируется стромальными клетками костного мозга. Кровь. 1997;90:3444–3455. [PubMed] [Google Scholar] 105. Сунгаран Р., Маркович Б., Чонг Б.Х. Локализация и регуляция экспрессии мРНК тромбопоэтина в почках, печени, костном мозге и селезенке человека с использованием гибридизации in situ.Кровь. 1997; 89: 101–107. [PubMed] [Google Scholar] 106. Кунисаки Ю. и др. Артериолярные ниши поддерживают гемопоэтические стволовые клетки в состоянии покоя. Природа. 2013 В прессе. Гистологическая характеристика подтипов сосудистых структур и доказательства того, что околоартериолярные мезенхимальные клетки сохраняют ГСК в состоянии покоя. [Бесплатная статья PMC] [PubMed] [Google Scholar]107. Пармар К., Маух П., Вергилио Дж. А., Сакштейн Р., Даун Д. Д. Распределение гемопоэтических стволовых клеток в костном мозге в зависимости от регионарной гипоксии. Proc Natl Acad Sci U S A.2007; 104: 5431–5436. [Бесплатная статья PMC] [PubMed] [Google Scholar]108. Такубо К. и др. Регуляция уровня HIF-1альфа необходима для гемопоэтических стволовых клеток. Клеточная стволовая клетка. 2010;7:391–402. doi: 10.1016/j.stem.2010.06.020. [PubMed] [CrossRef] [Google Scholar] 109. Вишнич Д. и соавт. Условная абляция линии остеобластов у трансгенных мышей Col2.3deltatk. Джей Боун Шахтер Рез. 2001;16:2222–2231. [PubMed] [Google Scholar] 110. Ву Дж.И. и др. Остеобластная регуляция В-лимфопоэза опосредована Gs{альфа}-зависимыми сигнальными путями.Труды Национальной академии наук Соединенных Штатов Америки. 2008; 105:16976–16981. doi: 10.1073/pnas.0802898105. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]111. Нагасава Т., Кикутани Х., Кисимото Т. Молекулярное клонирование и структура пре-В-клеточного фактора, стимулирующего рост. Труды Национальной академии наук США. 1994; 91: 2305–2309. [Бесплатная статья PMC] [PubMed] [Google Scholar]112. Не И, Хан YC, Цзоу YR. CXCR4 необходим для покоя примитивных гемопоэтических клеток.Журнал экспериментальной медицины. 2008; 205: 777–783. doi: 10.1084/jem.20072513. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]115. Химбург Х.А. и др. Плейотрофин регулирует удержание и самообновление гемопоэтических стволовых клеток в сосудистой нише костного мозга. Cell Rep. 2012; 2:964–975. doi: 10.1016/j.celrep.2012.09.002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]116. Смит-Бердан С. и соавт. Robo4 взаимодействует с CXCR4, чтобы определить локализацию гемопоэтических стволовых клеток в нишах костного мозга.Клеточная стволовая клетка. 2011; 8:72–83. doi: 10.1016/j.stem.2010.11.030. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]117. Смит-Бердан С., Шеперс К., Ли А., Пассег Э., Форсберг Э.С. Динамическая экспрессия лиганда Robo Slit2 в популяциях клеток костного мозга. Клеточный цикл. 2012; 11: 675–682. doi: 10.4161/cc.11.4.19146. [PubMed] [CrossRef] [Google Scholar] 118. Накамура-Исидзу А. и соавт. Белок внеклеточного матрикса тенасцин-С необходим в микроокружении костного мозга, предназначенном для гемопоэтической регенерации.Кровь. 2012;119:5429–5437. doi: 10.1182/blood-2011-11-393645. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]119. Стиер С. и др. Остеопонтин является компонентом ниши гемопоэтических стволовых клеток, который негативно регулирует размер пула стволовых клеток. Журнал экспериментальной медицины. 2005; 201:1781–1791. doi: 10.1084/jem.20041992. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]120. Нильссон С.К. и др. Остеопонтин, ключевой компонент ниши гемопоэтических стволовых клеток и регулятор примитивных гемопоэтических клеток-предшественников.Кровь. 2005; 106:1232–1239. doi: 10.1182/blood-2004-11-4422. [PubMed] [CrossRef] [Google Scholar] 121. Шеперс К. и др. Миелопролиферативная неоплазия ремоделирует эндостальную нишу костного мозга в самоусиливающуюся лейкемическую нишу. Клеточная стволовая клетка. 2013; 13: 285–299. doi: 10.1016/j.stem.2013.06.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]122. Уокли Ч.Р. и др. Индуцированный микроокружением миелопролиферативный синдром, вызванный гамма-дефицитом рецептора ретиноевой кислоты. Клетка. 2007; 129:1097–1110.[Бесплатная статья PMC] [PubMed] [Google Scholar]123. Уокли Ч.Р., Ши Дж.М., Симс Н.А., Пуртон Л.Е., Оркин С.Х. Rb регулирует взаимодействие между гемопоэтическими стволовыми клетками и их микроокружением костного мозга. Клетка. 2007; 129:1081–1095. [Бесплатная статья PMC] [PubMed] [Google Scholar]124. Киль М.Дж., Моррисон С.Дж. Неопределенность в нишах, поддерживающих гемопоэтические стволовые клетки. Нат Рев Иммунол. 2008; 8: 290–301. [PubMed] [Google Scholar]Дом гемопоэтических стволовых клеток
Хотя сосудистая ниша была обнаружена позже, чем остеобластная ниша, сосудистая ниша рассматривалась как альтернативная незаменимая ниша, управляющая регуляцией гемопоэтических стволовых клеток (ГСК).По мере значительного прогресса, достигнутого в нише этого типа, постепенно становится ясно, что основная работа сосудистой ниши берется на поддержку кроветворения. Однако по сравнению с тем, что было определено в механизмах, посредством которых остеобластная ниша регулирует гемопоэз, мы знаем меньше о сосудистой нише. В этом обзоре, основываясь на данных исследований, мы сосредоточимся на компонентной основе и различных функциях сосудистой ниши, которые обеспечивают нормальный процесс кроветворения в микроокружении костного мозга.Будут также обсуждаться возможные пути, выявленные в результате различных исследований, посредством которых эта среда выполняет свою функцию.
1. Введение
Гемопоэтические стволовые клетки человека (ГСК) сначала появляются у самого раннего эмбриона, затем перемещаются в печень и селезенку плода и, наконец, мигрируют в костный мозг (КМ). Самостоятельное обновление и дифференцировка ГСК — жестко регулируются как внутренними, так и внешними факторами клетки, и достигается пожизненный гомеостаз функциональных ГСК [1-3].Предполагается, что в области костного мозга должны существовать специфические микроокружения, содержащие ГСК и другие поддерживающие клетки, организующие взаимодействие между клетками и клетками, клетками и факторами для поддержания определенных аспектов кроветворения, таких как выживание ГСК, самообновление, и дифференциация. Эти процессы были связаны с рядом различных типов стромальных клеток и сигнальных путей. BM имеет гомогенную архитектуру, лишенную структурных и функциональных перегородок, а периферия полости взрослого BM содержит компартменты с различными свойствами [4].Недавнее исследование показало, что ГСК преимущественно локализуются в эндостальных зонах, где они наиболее тесно взаимодействуют с синусоидальными и несинусоидальными микрососудами костного мозга, формирующими своеобразную систему кровообращения [5]. Эти особые гемопоэтические микроокружения также называются «нишами», впервые предложенными Шофилдом в 1978 г. для описания областей, в которых находятся ГСК [6].
Согласно современной гипотезе, специализированная область внутри костного мозга, в которой обитают ГСК, подразделяется на два типа: остеобластная ниша и сосудистая ниша костного мозга [7, 8].HSCs в основном поддерживаются эндостальной нишей остеобластов, которая обеспечивает спокойное микроокружение HSC. А сосудистая ниша может регулировать пролиферацию, дифференцировку и мобилизацию ГСК [9]. Тем не менее, разработки в этой области будут полезны для продвижения экспансии и трансплантации ГСК в будущем.
2. Взаимодействие ГСК со специализированным микроокружением
Еще в начале 1970-х годов указывалось на локализацию недифференцированных гемопоэтических клеток вблизи эндостальной поверхности кости [10, 11], а увеличение числа остеобластов сопровождалось увеличением количества ГСК КМ in vivo [12].А последующие исследователи предложили понятие «ниша». Основной задачей, отягощенной остеобластной нишей, является поддержание долговременных ГСК (ДТ-ГСК), способных поддерживать кроветворение в течение месяцев или даже жизни в виде покоящихся или низкоциклирующих клеток [7, 13–15]. Выявлено, что увеличение количества клеток остеобластной выстилки в области трабекулярной кости коррелирует с увеличением количества ЛТ-ГСК [13]. Однако, поскольку поддержание ГСК является лишь головным звеном кроветворения, остальные звенья гемопоэтических процессов внутри костного мозга, такие как мобилизация, миграция и дифференцировка ГСК, также важны для исследования кроветворения.Вот почему недавно обнаруженная сосудистая ниша привлекает внимание исследователей к ее решающей роли в системе крови.
2.1. Модель ниши HSC
Исследования на Drosophila, C. elegans, и многих млекопитающих выявили критические особенности ниш стволовых клеток с помощью сигналов, исходящих от определенных клеток, и адгезии между стволовыми клетками и поддерживающими стромальными клетками или внеклеточным матриксом (ECM). ), которые обладают способностью управлять самообновлением, выживанием, поддержанием и дифференцировкой стволовых клеток [16, 17].Для выполнения таких функций комплексные компоненты ниш являются фундаментальными блоками, из которых строится такой «микродворец». Интегрируя данные из многочисленных систем, мы можем создать гипотетический «список деталей» для ниш HSC, включая сам HSC; стромальные клетки; белки ЕСМ; кровеносный сосуд; нейронные входы; и эндотелиальные клетки (ЭК) (рис. 1). Взаимодействия HSC-ниши влияют на самообновление [7, 13], состояние покоя [18-20] или мобилизацию HSCs [21], в зависимости от множества внутренних и внешних факторов.
2.2. Взаимодействие различных компонентов в нишах ГСК
Путем взаимодействия с этими «блоками» ниш ГСК клетки-предшественники, полученные из ГСК, расположенных на внутренней поверхности костного мозга, мигрируют в кровеносные сосуды в центре полости костного мозга и затем дифференцируются в зрелые клетки . Было высказано предположение, что HSC прикрепляются к поверхности остеобластов на основании наблюдения клеток N-кадгерина + в эндосте, которые экспрессируют некоторые маркеры HSC [13, 14, 22].Продуцируя регуляторы, такие как ангиопоэтин [14] и остеопонтин [18], они способны регулировать размер пула HSC in vivo . Остеокласты также являются важными компонентами, необходимыми для резорбции кости. Активация остеокластов также уменьшает компоненты ниши стволовых клеток, SDF-1 и SCF, что приводит к последующей мобилизации ГСК [23]. Форма кости регулируется остеобластами и остеокластами в динамическом равновесии в стационарных условиях [24, 25], влияя на судьбу ГСК.Более того, поскольку поверхность эндоста сильно васкуляризирована, сосудистые и периваскулярные клетки, такие как ретикулярные клетки, адипоциты и мезенхимальные предшественники, могут способствовать образованию ниш HSC в эндосте или рядом с ним [26, 27]. Наряду с остеобластной нишей этот тип микроокружения был определен как «сосудистая ниша» [8]. И дальнейший эксперимент подтвердил, что воздействие на HSCs эндотелиальных клеток (ECs), выделенных из разных тканей, приводит к различному росту HSC и способности к репопуляции [28].
Было показано, что периваскулярные CXCL12-обильные ретикулярные клетки (CAR-клетки) необходимы для поддержания содержания HSC BM in vivo [26]. Напротив, адипоциты костного мозга отрицательно влияют на размер и функцию пула ГСК [29]. Совсем недавно было продемонстрировано, что нестин-экспрессирующие МСК (NES + МСК) тесно связаны с поддержанием ГСК и могут размножать остеобласты в нише ГСК BM [30]. Таким образом, согласно тому, что было упомянуто выше, вполне вероятно, что совместная работа костных клеток, гемопоэтических клеток и ЭК в остеобластной нише регулирует гемопоэз.
3. Различные роли сосудистой ниши в гемопоэзе
Исследования сосудистой ниши совершили прорыв гораздо позже того, что было сделано для остеобластной ниши. Ясно лишь то, что посредством взаимодействия между ГСК и ЭК ГСК управляют процессами самообновления и дифференцировки [31–33]. Гематопоэтические клетки-предшественники, полученные из HSCs, находящихся в состоянии покоя в остеобластной нише, проникают в ECs, чтобы находиться в сосудистой нише, затем дифференцируются в различные типы клеток крови и в конечном итоге попадают в систему кровообращения [32, 34, 35] (Рис. 2).
3.1. Клеточная основа сосудистой ниши
Клеточная основа сосудистой ниши неоднородна по своей природе и происхождению: синусоидальные сосуды снабжаются артериолами и капиллярами, которые происходят от развилки артериальных сосудов, охватывающих полость костного мозга [32]. Синусоиды связаны межсинусоидальными капиллярами и в совокупности впадают в центральный синус. В дополнение к предоставлению ниши для самообновления, экспансии и поддержания HSCs, синусоидальные эндотелиальные клетки (SECs) играют роль как в обеспечении платформы для дифференцировки гемопоэтических клеток, так и в канале для мобилизации и возвращения гемопоэтических клеток в и из них. БМ [32, 36].Идея о том, что синусоиды могут представлять пролиферативную нишу, согласуется с недавними исследованиями, указывающими на то, что Е-селектин, молекула адгезии, конститутивно экспрессирующаяся в определенных синусоидальных микродоменах костного мозга, способствует пролиферации ГСК, а блокада Е-селектина защищает ГСК после химиотерапии или -облучения [37]. , 38]. Таким образом, прерывистые SEC составляют функциональную гемопоэтическую сосудистую нишу и играют важную роль в гемопоэтических процессах. Взаимодействуя с ГСК и окружающими их, сосудистая ниша поддерживает ряд гемопоэтических процессов при активации ГСК.
Гистохимический анализ молекулы активации сигнальных лимфоцитов (SLAM) показал, что около 60% SLAM-маркированных ГСК визуализируются вблизи СПК, что указывает на существование альтернативной ниши в сосудистой зоне [8, 39]. Было показано, что после трансплантации КМ ГСК предпочитали вживляться в сосудистые домены КМ у мышей, не получавших никакого другого лечения [40].
3.2. Функция сосудистой ниши
До сих пор было известно, что сосудистая ниша влияет на гемопоэз и поддержание ГСК.Во время кроветворения ГСК мобилизуются из покоящегося состояния [32, 41], мигрируют в кровоток, проникая через стенку синусоидов [35], и, наконец, дифференцируются в несколько видов клеток крови [42]. Как только BM находится в состоянии стресса, в селезенке или печени происходит экстрамедуллярный гемопоэз в сосудистой нише, чтобы дополнить функцию при аномальном BM [43]. В процессе участвуют различные факторы, которые будут обсуждаться ниже.
Кроме того, эндотелиальные клетки также играют важную роль во время кроветворной регенерации, как показали другие исследования.Например, трансплантация эндотелиальных клеток-предшественников ускоряла восстановление синусоидальных сосудов костного мозга после облучения, что коррелировало с более высоким восстановлением клеточности костного мозга и ГСК [44].
В селезенке мышей, мобилизованных циклофосфамидом/Г-КСФ, около 62% ГСК с отчетливым поверхностным маркером CD150 + CD48 − CD41 − наблюдались в контакте с синусоидальными ЭК, в то время как остальные клетки (38%) всегда находились вблизи синусоид [4]. В другом исследовании при тесном контакте с ЭК синусоидов печени происходит экстрамедуллярное печеночное кроветворение, что свидетельствует о том, что печень обеспечивает сходное микроокружение для поддержания и роста кроветворных клеток по сравнению с КМ [8, 45, 46].Все эти данные свидетельствуют о том, что сосудистая ниша может действовать как подкрепление для поддержки поддержания и дифференцировки HSCs, когда BM работает ненормально.
3.3. Взаимодействие с остеобластной нишей
Поскольку синусоидные ЭК действуют как альтернативная ниша для ГСК наряду с остеобластной нишей, было выдвинуто предположение, что сосудистая ниша подвергается дифференцировке ГСК, в то время как остеобластная ниша предлагает спокойное микроокружение [32]. Это можно объяснить тем фактом, что сосудистая ниша по сравнению с остеобластной нишей обеспечивает более богатую питательными веществами микросреду, характеризующуюся более высокими концентрациями кислорода и факторов роста, что приводит к тому, что ГСК являются пролиферативными и зрелыми клетками крови, в конечном итоге высвобождающимися в периферический кровоток [32]. , 41].Следуя этому процессу, другой вероятной функцией этого типа ниш является поддержка HSCs в трансэндотелиальной миграции. Клетки крови могут легко проходить через синусоидальную стенку, поскольку она состоит из одного слоя ЭК [35]. Следовательно, сосудистая ниша может не только способствовать пролиферации и дифференцировке ГСК при гематопоэтическом стрессе [42], но также обеспечивает немедленный выброс ГСК в кровоток.
Как правило, остеобластная ниша может поддерживать ГСК в состоянии покоя [14, 47]; между тем, сосудистая ниша регулирует мобилизацию HSCs.Однако это показало, что ECs, взаимодействующие с HSCs, могут играть необходимую роль в поддержании или пролиферации HSCs in vitro или in vivo . А так как многие HSCs в контакте с BMECs пролиферируют в любой момент времени, вероятно, что HSCs внутри сосудистого костного мозга являются самообновляющимися, а не долговременно бездействующими [8]. Брандт и его коллеги сообщили, что микрососудистые ЭК свиньи (PMVEC) могут опосредовать экспансию человеческих HSC ex vivo , а HSC, выделенные из совместной культуры PMVEC, были способны к репопуляции в BM мышей SCID [48]. In vivo гемопоэз хозяина может быть восстановлен у летально облученных мышей после трансплантации ЭК, выделенных из головного мозга или легких, что указывает на важную роль ЭК в самообновлении и репарации HSC [33]. Инфузии аллогенных эндотелиальных клеток-предшественников вызывали восстановление кроветворения и увеличение LT-HSCs у мышей с миелосупрессией, индуцированной облучением всего тела (TBI-), что позволяет предположить, что ECs положительно регулируют регенерацию HSC in vivo [44].
Кроме того, сообщалось, что гемопоэтические клетки могут происходить из ЭК. На раннем этапе развития наряду с васкуляризацией происходит кроветворение, и эндотелий отвечает за появление ГСК в области аорта-гонад-мезонефрос (АГМ) in vivo [49]. Кроме того, во многих исследованиях было показано, что некоторые клетки гемопоэтических кластеров, находящихся на дне дорсальной части аорты у видов от амфибий до человека, плотно контактируют с ЭК в нарушенной области просвета [49–51].Впоследствии с помощью экспериментов по отслеживанию клонов как у мышей, так и у рыбок данио стало ясно, что гемогенный эндотелий в вентральной части дорсальной аорты является первоначальным источником HSCs [52-56]. Таким образом, эти элегантные данные указывают на то, что активность большинства HSCs тесно связана с ECs, а эндотелий в AGM является первоначальным источником HSCs. Затем ГСК из АГМ дифференцируются в печени плода и, наконец, оседают во взрослой КМ [52, 57]. Таким образом, эндотелий работает как фабрика, сохраняя способность продуцировать ГСК на ранних стадиях развития организма.Вполне возможно, что эндотелий, выделенный из AGM, может вылечить гематологическое заболевание без присутствия имплантированных HSC.
4. Молекулярная основа для сосудистой ниши, регулирующей гемопоэз
Гематопоэз, регулируемый сосудистой нишей, относится к функциям, которые обсуждались выше, включая подвижность, трансэндотелиальную миграцию, гемопоэтическую дифференцировку и поддержание ГСК внутри или вне костного мозга. Также включен соответствующий компонент, окружающий HSC.Таким образом, остро стоит вопрос: через какой механизм или молекулярную основу этот тип ниши связывает ГСК и их окружение, чтобы поддерживать в порядке его регулярную работу по кроветворению? Молекулы, регулирующие гематопоэз, были обобщены в таблице 1.
| ||||||||||||||||||||||||||||||||||||||
4.1. Стромальный клеточный фактор-1 (SDF-1)
Было установлено, что стромальный клеточный фактор-1 (SDF-1), экспрессируемый ЭК КМ, может стимулировать ГСК и ЭК КМ, что приводит к усилению трансэндотелиальной стромальную миграцию за счет активации молекул адгезии [58, 59]. Процесс, при котором SDF-1 индуцирует трансэндотелиальную миграцию, может зависеть от иммобилизованной фракции хемокинов, связанной с компонентами внеклеточного матрикса (ECM) и стромальными клетками костного мозга, называемой «гаптотактическим градиентом» [60, 61].Как правило, покоящиеся HSC, застрявшие в нише остеобластов, должны быть сначала мобилизованы, если необходим гемопоэз. Имеются данные, показывающие, что SDF-1, экспрессируемый остеобластами, помогает HSC возвращаться в нишу остеобластов [62]. Однако экспрессия SDF-1 на ECs предвосхищает ключевую процедуру мобилизации HSCs.
Действительно, несколько сообщений указывают на то, что SDF-1 играет решающую роль в поддержании функции HSC, включая удержание в костном мозге, состояние покоя и репопуляционную активность.Однако также хорошо известно, что в костном мозге SDF-1 в основном экспрессируется тремя популяциями периваскулярных стромальных клеток: ретикулярными клетками с избытком SDF-1, нестин-стромальными клетками и стромальными клетками, содержащими рецептор лептина. Хотя эндотелиальные клетки костного мозга также экспрессируют SDF-1, делеция SDF-1 в клетках специфической ниши указывает на то, что эндотелиальные клетки вносят лишь незначительный вклад, а мезенхимальные предшественники являются основным источником SDF-1, который поддерживает HSCs. В соответствии с представлением о том, что множественные составляющие ниши выполняют разные функции, условная делеция Cxcl12 с помощью Lepr-credos не влияет на количество HSC в костном мозге, но индуцирует их мобилизацию [63].
In vitro , в присутствии SDF-1, экспрессируемого BMECs, фрагментация мегакариоцитов увеличивается, когда CXCR-4- (SDF-1 рецептор-) положительные мегакариоциты мигрируют через слой BMECs, предполагая, что контакт между мегакариоцитов и BMEC важен для тромбопоэза in vitro [36, 64]. Таким образом, SDF-1 способен повышать сродство и миграционную способность мегакариоцитов через BMEC. Кроме того, было показано, что эндотелиальный FGF- (фактор роста фибробластов-) 4 поддерживает адгезию мегакариоцитов к BMEC.Сообщалось, что SDF-1 и FGF-4 индуцируют экспрессию молекул адгезии, в том числе очень поздний антиген- (VLA-) 4, экспрессируемый мегакариоцитами, и VCAM-1, экспрессируемый BMEC [34, 65]. Аналогичным образом, антитело CXCR-4 останавливает восстановление тромбоцитоза после лечения 5-ФУ у мышей c-Mpl -/- и приводит к уменьшению мегакариоцитов, а также истощению сосудистой ниши [36], что демонстрирует, что SDF-1 играет роль ключевую роль в дифференцировке ГСК. Более того, SDF-1 может способствовать сосудисто-зависимому тромбопоэзу у мышей с дефицитом тромбопоэтина (ТПО) [36].Поскольку ЭК также маркируются SDF-1, возможность того, что механизм ЭК, направляющий самообновление HSCs, имеет отношение к SDF-1, остается, однако молекулярный подход находится в стадии изучения.
На основании механизма действия гранулоцитарного колониестимулирующего фактора (Г-КСФ) было высказано предположение, что градиентный сдвиг различных типов SDF-1 может вызывать мобилизацию ГСК. Мобилизация может быть усилена SDF-1 на ECs, способствуя проникновению гемопоэтических предшественников в эндотелий, что приводит к дифференцировке.Так, мобилизация, индуцированная G-CSF, сопровождается постепенным снижением остеобластического SDF-1 и повышением экспрессии CXCR- (endothelial SDF-1 рецептор-) 4 на ГСК [66]. По сходному механизму экспрессия Tie2 (эндотелиального рецептора ангиопоэтина-1 и ангиопоэтина-2) снижается в сосудистой нише костного мозга во время стационарного гемопоэза. HSC также маркируются Tie2, который обеспечивает адгезию HSC к остеобластам, регулируя состояние покоя в остеобластной нише [14]. И ингибирование передачи сигналов Tie2 на ECs приводило к нарушению восстановления сосудистой ниши, а также к замедлению восстановления гемопоэза [67].Таким образом, Tie2 что-то делает с поддержанием HSCs, а подавление Tie2 с помощью G-CSF направляет подвижность HSCs [14].
4.2. Notch Signaling
Более того, недавнее исследование показало, что ECs, заставляющие долговременные HSCs подвергаться потенциалу самообновления, имеют большое отношение к передаче сигналов Notch [68]. Эндотелиальные клетки экспрессируют лиганды Notch Jagged, которые поддерживают долговременную пролиферацию HSC и предотвращают их истощение. Экспрессия Notch в HSCs зависит от передачи сигналов Wnt и необходима для поддержания недифференцированного состояния [69].При использовании трансгенных мышей-репортеров Notch (TNR.GFP + ), у которых сигнальный путь Notch (экспрессируемый на ЭК) стимулирует экспрессию GFP, было подтверждено, что ЭК поддерживают долгосрочную экспансию TNR.Gfp + cKit + . Sca-1 + Lineage − (TNR.Gfp + KLS), но не Notch2 −/− Notch3 −/− CD34 − Flt-3 − KLS Кроме того, активация ангиогенной экспрессии notch-лигандов смещает баланс между экспансией и клон-специфичной дифференцировкой долгосрочных HSCs в сторону экспансии LT-HSCs, замедляя дифференцировку [68].Т.о., внутри сосудистой ниши высвобождение экспрессирующих ангиокрин лигандов notch с помощью ECs функционирует, чтобы установить инструктивную нишу для восстановления пула LT-HSC [45].
5. Сосудистые ЭК-секретирующие факторы, опосредующие самообновление и регенерацию ГСК
Выше упоминалось, что сосудистая ниша обладает способностью помогать ГСК самообновлению и репопуляции в КМ, в которых процессы ЭК играют решающую роль [33, 44]. Т.о., передача сигналов BM EC жизненно важна для восстановления кроветворения in vivo .Химбург и др. и Кобаяши и др. предположили, что растворимые факторы, продуцируемые ЭК BM, могут быть ответственны за самообновление и регенерацию HSC in vivo , но детальные механизмы остаются неизвестными [63, 70]. Механизм действия нескольких факторов будет рассмотрен ниже.
5.1. Фактор стволовых клеток (SCF)
Предполагается, что фактор стволовых клеток (SCF) экспрессируется эндотелиальными клетками, фибробластами костного мозга, остеобластами, периваскулярными стромальными клетками, экспрессирующими CXCL12, и мезенхимальными стволовыми клетками, экспрессирующими нестин [71, 72].Недавно сообщалось, что у мышей с делецией SCF, экспрессируемого на эндотелиальных клетках (Tie2-cre;), наблюдается снижение частоты LT-HSC с уменьшенной способностью к репопуляции во время трансплантации конгенного костного мозга. Делеция SCF в гемопоэтических клетках, остеобластах или клетках Nestin + не изменяет функции HSC, что идентифицирует SCF как специфическую молекулу in vivo, полученную из BM EC, которая регулирует HSC [1].
5.2. Эндотелиальные селектины (E-селектин)
Эндотелиальные селектины (E-селектин) представляют собой молекулы клеточной адгезии, которые экспрессируются в сосудистых нишах HSC, к которым прикрепляются гемопоэтические стволовые клетки и клетки-предшественники [37].Недавние исследования показали, что молекула адгезии Е-селектин экспрессируется исключительно эндотелиальными клетками костного мозга в сосудистой нише. У мышей с нокаутом E-селектина (Sele -/- ) было усилено состояние покоя HSC, а потенциал самообновления увеличился. При введении антагониста E-селектина результаты показали, что E-селектин способствует пролиферации HSC и является важным компонентом сосудистой ниши [37].
5.3. Гликопротеин 130 (gp130)
Гликопротеин 130 (gp130), сигнальная субъединица, общая для семейства рецепторов интерлейкина-6 (IL-6) цитокинов, также вносит значительный вклад в гемопоэз.Делеция gp130 в гематопоэтических и ЭК мышей с помощью рекомбинации генов, опосредованной Cre/ loxP , вызывала дисфункцию костного мозга, приводящую к тяжелой анемии во взрослом возрасте. Нормальный гемопоэз восстанавливался при трансплантации gp130-дефицитного BM облученным мышам дикого типа, тогда как гемопоэтические дефекты все еще существовали при трансплантации BM дикого типа облученным gp130-дефицитным мышам. Этот результат показал, что экспрессия gp130 на ECs, а не на самих гемопоэтических клетках, оказывает большое влияние на гемопоэз, предоставляя доказательства того, что ECs зависят от передачи сигналов gp130 для поддержки кроветворения, реагируя на сигналы от цитокинов семейства IL-6 [73].
5.4. Плейотрофин (PTN)
Недавно было показано, что гепарин-связывающий фактор роста, плейотрофин (PTN), секретируемый ЭК КМ, вызывает регенерацию ГСК у мышей после облучения всего тела высокой дозой (ЧМТ). Эти эффекты можно объяснить связыванием и ингибированием белкового рецептора тирозинфосфатазы-зета (PTPRz) на ГСК КМ [70].
6. Заключение
Сосудистая ниша играет ключевую роль в поддержании гемопоэза, включая подвижность, трансэндотелиальную миграцию и гемопоэтическую дифференцировку.Более того, по сравнению с длительно спящими ГСК в остеобластной нише, сосудистая ниша поддерживает самообновление ГСК, что приводит к поддержанию ГСК. Таким образом, сосудистая ниша может регулировать гемопоэз и поддерживать самообновление HSCs. Таким образом, воссоздание этой ниши в организме с нарушением кроветворения может заменить терапевтический метод трансплантации ГСК. Эндотелиальные терапевтические методы прольют свет на заболевания кроветворения, такие как апластическая анемия и лейкемия, ускорят восстановление кроветворения после химиотерапии или сократят время реконструкции кроветворения после трансплантации костного мозга.Хотя было показано, что механистические пути, такие как передача сигналов Notch и CXCR-4-SDF-1, способствуют эндостальной и ретикулярной клеточной регуляции HSCs судьбы in vivo , механизмы, с помощью которых сосудистая ниша регулирует поддержание и регенерацию HSCs, остаются менее четко определенными. . И выяснение механистических взаимодействий и корегуляции кроветворения остеобластами, ECs и другими клетками микроокружения BM будет центральной задачей ближайшего десятилетия. Дальнейшие исследования продвинут существующие результаты вперед и сделают большой прорыв в сосудистой нише.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в связи с публикацией данной статьи.
Вклад авторов
Ниннин Хэ и Лу Чжан внесли равный вклад в настоящую работу.
Благодарности
Эта работа была поддержана Национальной ключевой научной программой Китая (2011CB964903), Национальным фондом естественных наук Китая (81371620), Тяньцзиньским фондом естественных наук (12JCZDJC24900) и Программой для выдающихся талантов нового века в университете (NCET). Государственное министерство образования Китая (NCET-12-0282).
Старая ниша костного мозга сдерживает омоложенные гемопоэтические стволовые клетки — Guidi — 2021 — СТВОЛОВЫЕ КЛЕТКИ
1 ВВЕДЕНИЕ
Старые гемопоэтические стволовые клетки (ГСК) способствуют развитию лейкемии, связанной со старением, и ремоделированию иммунной системы, связанному со старением. 1, 2 Старение приводит к увеличению количества ГСК в костном мозге (КМ), в то время как старые ГСК демонстрируют сниженный потенциал восстановления, миелоидную асимметрию, измененные профили экспрессии генов и эпигенетические модификации 3-7 и переключение от асимметричных до симметричных отделов. 8-10 При старении также отмечается увеличение частоты ГСК с неполярным распределением белков полярности. 11, 12 Старение HSC может даже повлиять на продолжительность жизни. 13 Механизмы, присущие HSC, а также внешние сигналы (ниша/микроокружение) могут вызывать старение HSC. 14, 15 Например, снижение связанной со старением повышенной активности малой RhoGTPase Cdc42 в старых HSC ex vivo с помощью специфического ингибитора активности Cdc42 CASIN 16 приводит к омоложению функции старых HSC.Эта юношеская функция старых омоложенных ГСК сохраняется при их трансплантации молодым реципиентам. 2, 9, 17
Молодые HSCs при трансплантации в старую нишу BM демонстрируют признаки старых HSCs. 18, 19 Это указывает на сильное влияние стареющей ниши на функцию ГСК. В соответствии с этой концепцией мы недавно продемонстрировали, что снижение уровня секретируемого цитокина остеопонтина (ОПН) в нише старого костного мозга придает признаки старения молодым ГСК, 19 , подобно увеличению размера пула ГСК (см. также Справку). 20), снижение частоты ГСК, полярных для белков полярности, и увеличение потенциала миелоидной дифференцировки ГСК. 19 Экспрессия OPN в BM ограничена эндостальной поверхностью кости. 21 Интересно, что секретируемый OPN в конечном итоге передает сигналы через интегрины α9β1 на HSC, чтобы регулировать активность Cdc42 и, таким образом, полярность HSC. 19, 22 Влияние ниши (молодые или старые) на омоложенные старые ГСК, следовательно, является критическим вопросом для разработки подходов к замедлению старения ГСК in vivo. Таким образом, целью этого исследования было определить, может ли старая ниша влиять на молодоподобную функцию омоложенных старых HSC.
2 МЕТОДЫ
2.1 Мыши
молодых мышей C57BL/6 (возраст 8-10 недель, Janvier, St. Berthevin Cedex, Франция). Старые мыши C57BL/6 (19-24 месяца) и мыши с нокаутом OPN (KO) (C57BL/6, 8-12 недель) были получены из внутреннего поголовья. C57BL/6.SJL- Ptprc a /Boy (BoyJ) молодых (8-10 недель) и старых (19-24 месяцев) мышей получали из Charles River Laboratories, Зульцфельд, Германия, или из внутренних склад.
2.2 Конкурентная трансплантация ДТ-ГСК
Двести старых LT-HSC (Ly5.1 + ) культивировали в течение 16 часов в HBSS + 10% FBS с 5 мкМ CASIN или без него при 37°C, 5% CO 2 , 3% O 2 и трансплантировали вместе с 3 × 10 5 клеток BM от молодых мышей C57BL/6 (Ly5.2 + ) в молодых, старых и молодых мышей-реципиентов OPN KO C57BL/6 (Ly5.2 + ). Донорский химеризм в крови и костном мозге анализировали до 23 недель после трансплантации.Для экспериментов, показанных на дополнительном рисунке 1, 200 молодых или старых LT-HSC (Ly5.1 + ) культивировали в течение 16 часов в HBSS + 10% FBS с 5 мкМ CASIN или без него при 37 °C, 5% CO 2 , 3% O 2 и трансплантированы вместе с 3 × 10 5 клетками костного мозга молодых мышей C57BL/6 (Ly5.2 + ) реципиенту C57BL/6 молодых (молодых в молодых) или старых (старых в старых) реципиентов мыши (Ly5.2 + ).
2.3 Проточная цитометрия и сортировка клеток
Иммуноокрашивание периферической крови (PB) и клеток костного мозга выполняли в соответствии со стандартными процедурами и анализировали на проточном цитометре LSRII или BD FACSAria III (BD BioSciences, Гейдельберг, Германия).Антитела: анти-Ly5.2 (клон 104), анти-Ly5.1 (клон А20), анти-CD3e (клон 145-2С11), анти-В220 (клон RA3-6B2), анти-Mac-1 (клон М1 /70) и анти-Gr-1 (клон RC57BL/6-8C5), анти-CD11b (клон M1/70), анти-B220 (клон RA3-6B2), анти-CD5 (клон 53-7.3), анти- Gr-1 (клон RB6-8C5), анти-Ter119 и анти-CD8a (клон 53-6.7). После истощения линии с помощью магнитной сепарации (Dynabeads, Invitrogen, Dreieich, Германия) клетки окрашивали анти-Sca-1 (клон D7), анти-c-Kit (клон 2B8), анти-CD34 (клон RAM34), анти-CD127. (клон A7R34), анти-CD16/CD32 (клон 2.4G2) (BD Biosciences, Гейдельберг, Германия), анти-Flk-2 (клон A2F10) и стрептавидин. Все антитела были получены от eBioscience (Dreieich, Германия), если не указано иное. Данные анализа FACS (стволовые клетки и клетки-предшественники) были нанесены на график в процентах от долгосрочных гемопоэтических стволовых клеток (LT-HSCs, гейтированных как LSK CD34 -/low Flk2 — ), краткосрочных гемопоэтических стволовых клеток (ST- HSCs, гейтированные как LSK CD34 + Flk2 − ), и мультипотентные предшественники (MPPs, гейтированные как LSK CD34 + Flk2 + ) среди донорских LSK (Lin neg c-kit) sca-1 + клеток).Обычные миелоидные предшественники (CMPS, закрыты как Lin NEG C-Kit + CD34 + CD16 / 32 — ), MegakaryoCyte-эритроциты (MEPS Gated как Lin Neg C-Kit + CD34 − CD16/32 − ) и гранулоцитарно-макрофагальных предшественников (GMPs, закрытые как Lin neg c-kit + CD34 + CD16/32 + ). Данные анализа FACS (дифференцированные клетки) представлены в виде процента В-клеток (B220+), Т-клеток (CD3+) и миелоидных (Gr-1+, Mac-1+ и Gr-1+Mac-1+) клеток среди донорский Ly5.1 + ячейки.
2.4 Иммунофлуоресцентное окрашивание
LT-HSC высевали на покровные стекла, покрытые фибронектином, инкубировали в течение 16 часов в HBSS, 10% FBS, 37°C, 5% CO 2 , 3% O 2 , с 5 мкМ CASIN, фиксировали 4% PFA. , промывали PBS, пермеабилизировали 0,2% Triton X-100 (Sigma, Дармштадт, Германия) в PBS в течение 20 минут и блокировали 10% ослиной сывороткой (Sigma, Дармштадт, Германия) в течение 30 минут. Антитело к α-тубулину (Abcam, Берлин, Германия, крысиное моноклональное ab6160), антитело к крысиному DyLight488, конъюгированное с DyLight488 (Jackson ImmunoResearch, Гамбург, Германия) и анти-h5K16ac (07-329) от Millipore, Дармштадт, Германия, анти- — кроличьи антитела, конъюгированные с DyLight549 (Jackson ImmunoResearch, Гамбург, Германия), в течение 1 часа при комнатной температуре.Предметные стекла наносили на реагент ProLong Gold Antifade с DAPI (Invitrogen, Molecular Probes, Dreieich, Германия), визуализировали на AxioObserver Z1 (Zeiss, Оберкохен, Германия) и анализировали с помощью программного обеспечения AxioVision 4.6.
2.5 Статистический анализ
Однофакторный дисперсионный анализ или двухфакторный дисперсионный анализ использовались для сравнения средних значений среди трех или более независимых групп. Последующий тест Бонферрони для сравнения всех пар набора данных был определен, когда общее значение P было <.05. Все статистические анализы проводились с помощью Prism версии 8.0.1.
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В предыдущих экспериментах молодые или старые Ly5.2 HSC трансплантировали молодым реципиентам Ly5.1. Аллель Ly5.1 у конгенных животных Ly5.1 содержит помимо Ly5.1 другие гены, которые могут влиять на кроветворение. 23 Новые эксперименты, в которых молодые Ly5.1+ HSC трансплантировали молодым Ly5.2+ реципиентам, а старые Ly5.1+ HSC трансплантировали старым Ly5.Реципиенты 2+ подтвердили «возрастную функцию» также старых Ly5.1+ HSCs при трансплантации старым Ly5.2+ реципиентам (дополнительная фигура 1), аналогично тому, что сообщалось ранее для старых Ly5.2 HSCs, трансплантированных молодым Ly5.1+ получатели. 17, 24 Приживление донорских клеток было (как и ожидалось) высоким в трансплантатах Ly5.1+ Y→Ly5.2+ Y (молодые в молодые, Y→Y) и низким в Ly5.1+ O→Ly5.2+ A (от старых к старым, O➔A) трансплантатам с более низким вкладом старых Ly5.1+ HSC в B- и T-клетки в PB (дополнительная фигура 1B, C).Мы также наблюдали более высокий вклад старых HSC в частоту миелоидных клеток в PB и в частоту LT-HSC и ST-HSC в BM (дополнительная фигура 1D-F). Кроме того, частота MPP в BM была снижена в O➔A по сравнению с трансплантатами Y➔Y (дополнительная фигура 1G), тогда как частоты CMP, GMP и MEP были одинаковыми в обеих экспериментальных группах (дополнительная фигура 1H-J) и таким образом, не подвержены старению. Недавно мы описали сниженную частоту HSCs, полярных для распределения тубулина, как новый признак среди старых HSCs.HSC от пожилых реципиентов, которым трансплантировали старые Ly5.1+ HSC, также показали низкую частоту HSC, полярных для тубулина (дополнительная фигура 1K, L). Конгенный интервал, который окружает локус Ly5.1, таким образом, не по-разному влияет на связанные со старением изменения фенотипов и функции старых Ly5.1+ HSCs по сравнению со старыми Ly5.2+ HSCs. Результат этих критических контрольных экспериментов позволил провести сравнительный анализ функции старых Ly5.1+ HSC в различных типах ниш в следующих экспериментах по трансплантации.
Для определения того, в какой степени возраст ниши влияет на молодоподобную функцию и полярность омоложенных ex vivo старых HSC, старых или старых HSC (Ly5.1+), подвергшихся воздействию ингибитора активности Cdc42 CASIN (омоложенные старые HSC 9 ) были трансплантированы вместе с клетками-конкурентами Ly5.2+ BM молодым (как опубликовано ранее 9 ) или в этих наборах экспериментов теперь также реципиентам в возрасте (рис. 1A). У старых мышей уровень цитокина OPN сильно снижен в резидентных остеобластах BM, тогда как молодая ниша, в которой отсутствует OPN, превращает молодые HSC в функционально старые в отношении фенотипов и функции HSC.Таким образом, 19, 25 Старые HSC и омоложенные старые HSC также трансплантировали реципиентам OPN KO, чтобы проверить, влияет ли отсутствие OPN в нише на функцию старых омоложенных HSC (рис. 1A). Донорский химеризм в крови и костном мозге анализировали до 23 недель после трансплантации.
Возрастная микросреда сдерживает CASIN-зависимое омоложение старых гемопоэтических стволовых клеток (ГСК). А, схематическое изображение экспериментальной установки: (Ly5.1+) 200 клеток HSC от старых доноров обрабатывали +/- CASIN в течение 16 часов ex vivo, а затем конкурентно трансплантировали либо молодому (Y), старому (A), либо молодому (OPN) реципиенту с нокаутом остеопонтина (KO) (Ly5. 2+) мыши. B, частота вклада старых доноров (клетки Ly5.1+) +/- CASIN в общее количество лейкоцитов в периферической крови (PB) молодых, старых и молодых мышей-реципиентов OPN KO (Ly5.2+). C, Частота старых В-клеток +/- CASIN. D, старые Т-клетки +/- CASIN. E, старые миелоидные клетки +/- CASIN среди донорских клеток Ly5.1+ в PB молодых, старых и молодых реципиентов OPN KO (Ly5.2+) мыши. F, частота старых LT-HSC +/- CASIN в костном мозге (BM) среди донорских клеток Ly5.1+ LSK у молодых, старых и молодых мышей-реципиентов OPN KO (Ly5.2+). Данные основаны на пяти экспериментальных повторах с тремя мышами-реципиентами на группу (например, n = 11-15 на группу). G, Частота встречаемости старых LT-HSC +/- CASIN донорского происхождения, поляризованных по тубулину, отобранных из разных экспериментальных групп (Ly5.2+) мышей через 23 недели после трансплантации. п = 5; ~ 40 клеток на образец в каждой экспериментальной повторности.H, репрезентативное распределение тубулина (зеленый) в старых донорских LT-HSC +/- CASIN (клетки Ly5.1+), отобранных из разных экспериментальных групп через 23 недели после трансплантации. Масштабная линейка = 6 мкм. Показаны средние значения + 1 SEM. * P < .05, *** P < .001
Уровень химеризма, поддерживаемый старыми или старыми омоложенными ГСК при трансплантации молодым, пожилым или реципиентам ОПН КО, был одинаковым для каждого типа донорских ГСК, за исключением старых ГСК, трансплантированных молодым реципиентам.В этих условиях, как уже сообщалось в 19 , старые HSC показали значительно более высокий химеризм по сравнению со старыми HSC, трансплантированными старым реципиентам. Не было дифференциального химеризма среди молодых, старых реципиентов или реципиентов OPN KO, которым трансплантировали старые омоложенные (обработанные CASIN) HSC (рис. 1B).
ЧастотаB-клеток у реципиентов, которым были введены старые или старые омоложенные HSC, была одинаковой у молодых, старых или реципиентов OPN KO (рис. 1C). Частота Т-клеток, полученных из старых омоложенных ГСК, была увеличена по сравнению с частотой у молодых реципиентов, которым трансплантировали старые ГСК.Эта частота была снижена у пожилых реципиентов или реципиентов OPN KO (рис. 1D). Это показывает, что юношеская частота Т-клеток (см. также дополнительную фигуру 1C), поддерживаемая старыми омоложенными HSC, на самом деле требует молодой ниши, и это не зависит от возраста тимуса (см. также ссылку 2) и что отсутствие OPN в ниши было достаточно, чтобы нарушить частоту Т-клеток, поддерживаемую старыми омоложенными HSC. Частота донорских миелоидных клеток (в % от донорского химеризма) была низкой у молодых реципиентов, которым трансплантировали старые омоложенные ГСК (рис. 1Е и ссылка 17).Старые омоложенные HSC при трансплантации старым или молодым реципиентам OPN KO снова поддерживали высокий уровень миелоидных клеток, аналогичный уровню, обнаруживаемому у молодых реципиентов, которым трансплантировали старые HSC (рис. 1E). Следовательно, старая ниша также оказывает сильное регулирующее влияние на уровень миелоидных клеток на периферии, независимо от возраста ГСК, аналогично тому, что мы наблюдали для Т-клеток.
Молодые реципиенты, получившие старые омоложенные HSC, показали низкую частоту LT-HSCs в пределах BM 17 (рис. 1F).У старых животных или животных с нокаутом OPN при трансплантации старых омоложенных HSC наблюдалась лишь слегка повышенная частота LT-HSC (рис. 1F). Старые HSC являются аполярными для распределения тубулина или маркера эпигенетической полярности h5K16ac, тогда как молодые или старые омоложенные HSC при трансплантации молодым реципиентам остаются более полярными для распределения тубулина или h5K16ac 17, 19 (рис. 1G,H). . Старые омоложенные HSC при трансплантации пожилым реципиентам показали низкую частоту клеток, поляризованных для тубулина, которая была аналогична таковой у старых HSC (рис. 1G, H; дополнительный рисунок 1K, L).Интересно, что HSC от реципиентов OPN KO, которым трансплантировали старые омоложенные HSC, имели частоту HSC, полярную для распределения тубулина, которая была более похожа на частоту, обнаруженную у молодых реципиентов (рис. 1G, H). Напротив, HSC от пожилых реципиентов, которым трансплантировали старые омоложенные HSC, показали частоту клеток, поляризованных по маркеру эпигенетической полярности h5K16ac, аналогичную частоте, обнаруживаемой у молодых реципиентов или реципиентов OPN KO (дополнительная фигура 1M). Эти результаты подразумевают, что хотя возрастная ниша влияет на уровень полярности тубулина, но не на полярность h5K16ac, этот эффект может быть не просто следствием низкого уровня OPN, как это наблюдается для дифференцировки Т-клеток (рис. 1D).
Старение приводит к снижению частоты MPP в BM, но не влияет на частоту CMP, GMP и MEP 17, 24 (дополнительный рисунок 1G-J). Мы наблюдали небольшое, но значительное увеличение частоты ST-HSCs у молодых реципиентов, которым вводили старые омоложенные HSC, по сравнению с молодыми реципиентами, которым вводили старые HSC (рис. 2A). Интересно, что на частоту MPP не влияло ни омоложение HSC, ни тип животного-реципиента (рис. 2B), ни какой-либо из перечисленных выше других типов клеток-предшественников в BM (дополнительный рисунок 1N-P).
Старая микросреда не повреждает мультипотентные клетки-предшественники, происходящие из старых омоложенных гемопоэтических стволовых клеток (ГСК). A, частота ST-HSC и B, частота мультипотентных предшественников (MPP) среди донорских клеток Ly5.1+ LSK в костном мозге (BM) молодых, старых и молодых реципиентов с нокаутом остеопонтина (OPN) (KO) ( Ly5.2+) мышей. C, Young-подобные фенотипы омоложенных старых HSCs, ослабленных стареющей нишей. Показаны средние значения + 1 SEM.* P < .05
4 ЗАКЛЮЧЕНИЕ
Наши данные демонстрируют, что устойчивая юношеская функция омоложенных старых HSC при трансплантации молодым реципиентам является, по крайней мере, частично комбинацией внутреннего и внешнего вклада стволовых клеток (ниша/микроокружение). Некоторые из признаков старения HSC, которые возвращаются в юношеское состояние, когда омоложенные старые HSC трансплантируют молодым мышам, возвращаются к старому фенотипу, когда омоложенные старые HSC находятся в состарившейся нише (рис. 2C).Для некоторых фенотипов это может быть связано со сниженным уровнем OPN в возрастных нишах. Таким образом, влияние ниши на омоложенные HSCs необходимо учитывать для подходов, направленных на омоложение старых HSCs in vivo.
БЛАГОДАРНОСТИ
Мы благодарим Cores в Ульмском университете и CCHMC за поддержку сортировки клеток и Tierforschungszentrum Университета Ульма за поддержку нашей работы с животными. Работа с животными была одобрена Regierungspräsidium Tübingen.Эта работа была поддержана грантами Deutsche Forschungsgemeinschaft (SFB 1279) Х. Гейгеру. Мы благодарим Люси Лио за любезный подарок нокаутных мышей OPN. Финансирование открытого доступа разрешено и организовано Projekt DEAL.
КОНФЛИКТ ИНТЕРЕСОВ
Авторы заявили об отсутствии потенциального конфликта интересов.
АВТОРСКИЕ ВКЛАДЫ
NG: концепция и дизайн, сбор и сбор данных, анализ и интерпретация данных, написание рукописи, окончательное утверждение рукописи; Г.М., В.С.: сбор данных; Ю.З.: интерпретация данных; M.C.F.: концепция и дизайн, интерпретация данных; HG: концепция и дизайн, финансовая поддержка, административная поддержка, предоставление учебного материала, интерпретация данных, написание рукописи, окончательное утверждение рукописи.
Молекулярный переключатель костномозгового происхождения между остеогенезом и кроветворением
- Марта Галан-Диез и
- Ставрула Куштень
- Кафедра физиологии и клеточной биофизики, Колледж врачей и хирургов, Колумбийский университет, Нью-Йорк, Нью-Йорк 10032, США
- Автор, ответственный за переписку: sk2836{at}cumc.columbia.edu
Аннотация
Гематопоэтические стволовые клетки (ГСК) находятся и поддерживаются в специализированных микросредах костного мозга, известных как ниши, состоящие из различных типов клеток. Среди них лептиновый рецептор (LepR), экспрессирующий хемокиновый лиганд CXC 12 (CXCL12), обильный Известно, что ретикулярные (CAR) клетки создают нишу для ГСК и в то же время дают начало остеобластам.Эти две функции клеток CAR/LepR + , по-видимому, строго, но обратно регулируются, чтобы обеспечить адекватное физическое пространство для HSC. Однако как остеогенез предотвращается в клетках CAR, чтобы сохранить пространство, доступное для HSCs, и гемопоэз остается неясным. В этом выпуске Genes & Development Сейке и его коллеги (стр. 359–372) сообщают, что транскрипционный фактор раннего В-клеточного фактора ( Ebf3 ) преимущественно экспрессируется клетками CAR/LepR + и ингибирует клетки CAR. дифференцировку в остеобласты с одновременным сохранением самообновления клеток CAR/LepR + .Используя условно нокаутные и ретровирусные системы, исследователи показали, что потеря Ebf3 в клетках CAR снижает количество HSC и приводит к остеосклерозу. Это исследование дает новое понимание требований к транскрипции. для формирования кости CAR-клетками путем идентификации Ebf3 как нишевого фактора, секретируемого клетками CAR/Lepr + , который регулирует взаимодействие между остеогенезом и гемопоэзом.
Одной из важнейших и уникальных функций скелета является обеспечение анатомических пространств для поддержания кроветворения.Костный мозг является основным местом пребывания гемопоэтических стволовых клеток (ГСК), где они самообновляются, пролиферируют или становятся постепенно ограничивается несколькими зафиксированными предшественниками и/или отдельными линиями, дающими начало различным типам зрелых кровяные клетки. В костном мозге ГСК динамически взаимодействуют со сложной сетью стромальных клеток. Такие группы клеток представляют собой совокупность нескольких специализированных «микрониш», которые поддерживают кроветворение, влияя на различные функции HSC, такие как самонаведение, мобилизация, покой, самообновление или приверженность родословной.Поучительные подсказки предоставляются как необходимые аутокринные, эндокринные и паракринные сигналы, а также прямые межклеточные взаимодействия между стромальными нишами и HSCs.
Многочисленные исследования изучали роль линии остеобластов в ГСК и указывают на то, что регуляция остеобластов гемопоэза зависит от стадии дифференцировки остеобластов (Sacchetti et al. 2007; Wu et al. 2008; Méndez-Ferrer et al.2010 г.; Кальви и др. 2012 г.; Кунисаки и др. 2013). В то время как наиболее незрелая подгруппа некоммитированных предшественников влияет на поддержание и пролиферацию HSC, коммитированные предшественники или зрелые остеобласты приспосабливают дифференцировку HSC вдоль лимфоидных, миелоидных и эритроидных линий (Calvi et al. 2003; Zhu et al. 2007; Ding and Morrison 2013; Krevvata et al. 2014). В самом деле, самообновляющиеся остеопредшественники клеток в костном мозге могут образовывать поддерживающие ниши HSC (Sacchetti et al. 2007). Мезенхимальные стволовые клетки (МСК) эндостальной ниши в непосредственной близости от ГСК экспрессируют секретируемые молекулы или молекулы клеточной поверхности. которые регулируют состояние покоя HSC (Nilsson et al.2005 г.; Зильберштейн и др. 2016). Нестин-позитивные МСК продуцируют растворимые факторы, участвующие в поддержании ГСК, такие как хемокиновый лиганд 12 СХС (CXCL12) и фактор стволовых клеток (SCF) и колокализуются с HSCs, а их истощение приводит к мобилизации HSC (Méndez-Ferrer et al. 2010). Эти наблюдения показали, что остеогенез и поддержание HSC в костном мозге могут быть обратно пропорциональны.
Периваскулярные ниши регулируют поддержание и покой HSC.Артериолярная ниша, отмеченная редкими NG2 (CSPG4 + ) перицитами, ингибирует цикличность HSC и снижает функциональные долгосрочные репопуляции HSC в костном мозге (Kunisaki et al. 2013). Отдельная перисинусоидная ниша, содержащая большинство делящихся и неделящихся HSC, по-видимому, регулирует взаимодействие. между остеогенезом и кроветворением. Клетки в перисинусоидной нише, которые экспрессируют высокие уровни CXCL12 и SCF (CXCL12-богатые ретикулярные [CAR] клетки) составляют большинство периваскулярных клеток (Omatsu et al.2010) и перекрываются с перисинусоидальными клетками, идентифицированными на основании их экспрессии рецептора лептина (LepR) (Zhou et al. 2014). CAR и CAR/LepR + МСК, составляющие ~0,3% клеток костного мозга, включают почти все фибробласты, образующие колонии (КОЕ-Ф), являются основным источником адипоцитов и остеобластов в костном мозге взрослых мышей и необходимы для поддержки поддержания HSC. Тем не мение, большинство клеток CAR/LepR + могут оставаться недифференцированными в полости костного мозга, и остается неясным, как предотвращается остеогенез в большинстве случаев. CAR/LepR + клеток для поддержания пространства, доступного для HSCs и гемопоэза.
Чтобы ответить на этот вопрос, Seike et al. (2018) стремились идентифицировать регуляторы транскрипции клеток CAR и сосредоточились на факторах транскрипции раннего В-клеточного фактора. (Эбф) семья. Они обнаружили, что Ebf3 преимущественно экспрессируется в клетках CAR по сравнению с эндотелиальными, гемопоэтическими и стромальными клетками взрослого костного мозга. Отслеживание линии клеток Ebf3 , экспрессирующих CAR/LepR + , проводили с использованием тройных мутантов мышей, экспрессирующих CreERT2 , нокаутированных в эндогенном локусе Ebf3 , трансгене Cxcl12-GFP и Cre-активированных мышах, экспрессирующих CreERT2 Помидор (tdTomato).Количество Ebf3 -экспрессирующих CAR-клеток, дающих начало адипоцитам и остеобластам в костном мозге, оставалось стабильным в течение курса лечения. 13 мес., предполагая, что они способны к самообновлению. Условная делеция Ebf3 в клетках CAR/LepR + или во всех МСК развивающихся конечностей приводила к уменьшению числа долговременно репопулирующих ГСК (ДТ-ГСК), мегакариоцитов/эритроцитов. предшественники (MEP) проэритробласты, общие лимфоидные предшественники (CLP) и предшественники гранулоцитов/макрофагов (GMP).Кроме того, у старых мышей количество функциональных ГСК уменьшалось. Этот скомпрометированный гемопоэтический фенотип коррелирует с повышенным количества остеобластов и остеогенеза и более низкой экспрессии Cxcl12 и Scf в клетках CAR/LepR + . В результате у старых мышей развился остеосклероз. Интересно, что остаточный бета-положительный рецептор тромбоцитарного фактора роста (PDGFRβ + ) Sca-1 — CD31 — CD45 — и Ter119 — CAR клетки имели гораздо более высокие уровни экспрессии маркеров остеобластов.В совокупности эти результаты показали, что кроветворная нишевая функция стволовых клеток и клеток-предшественников (HSPC) клеток CAR была серьезно нарушена, и что большинство клеток CAR было более дифференцировались в линию остеобластов в отсутствие Ebf3 .
Чтобы лишить клетки CAR всех экспрессированных изоформ Ebf , исследователи инактивировали как Ebf3 , так и Ebf1 в МСК, включая клетки CAR.В возрасте 1 недели в костном мозге образовались полости, а количество CAR-клеток не изменилось. нормальная морфология с длинными отростками; однако у них была увеличена экспрессия маркеров остеобластов Osterix , Col1a1 и Osteocalcin и снижена экспрессия Cxcl12 и Scf по сравнению с контрольными животными. Это свидетельствует о том, что большинство клеток CAR более дифференцированы в остеобласты. линии у мутантных животных.Общее количество гемопоэтических клеток и количество LT-HSC, MEP, проэритробластов, CLP и GMP. действительно были снижены у мутантов. К 26-недельному возрасту у мышей с двойным нокаутом наблюдалась окклюзия полости костного мозга и снижение Сотовый номер автомобиля. Следовательно, Ebf1/Ebf3 , по-видимому, не определяют судьбу клеток CAR во время эмбриогенеза, но необходимы для ингибирования дифференцировки клеток CAR в линия остеобластов для поддержания пространств костного мозга.
Чтобы изучить соответствующие вклады Ebf3 и Ebf1 в поддержание HSC и дифференцировку остеобластов, исследователи изучали скелетные и гемопоэтические фенотипы взрослых CAR/LepR + клеточно-специфические Ebf1 -/- ;Ebf3 +/- и Ebf1 + 5 ; Ebf3 -/- мышей.Серьезная редукция предшественников ГСК в костном мозге приводила к экстрамедуллярному кроветворению и заметному увеличению в трабекулярной костной массе. Это было связано с увеличением числа остеобластов без изменений в остеокластах, что наблюдалось в Ebf1 -/+ ; Ebf3 -/- мышей, но не у Ebf1 -/- ;Ebf3 +/- мышей в возрасте 34 недель. клетки CAR из CAR/LepR + клеточно-специфические Ebf1 -/+ ; Ebf3 -/- мышей, но не Ebf1 -/- ;Ebf3 +/- мышей, имели более высокую экспрессию маркеров остеобластов.Следовательно, активности одного функционального аллеля Ebf3 достаточно для поддержания костномозговых пространств и ГСК и подавления остеобластогенеза, тогда как одного аллеля Ebf1 недостаточно. Наконец, сверхэкспрессия Ebf1 и Ebf3 в отсортированных CAR-клетках увеличивала экспрессию Cxcl12 и Scf и снижала их остеогенный потенциал в культуре. Этот эффект зависел от ДНК-связывающего домена Ebf3 .
В заключение, это исследование идентифицирует Ebf3 как молекулярный триггер, который действует как клеточно-автономным, так и клеточно-неавтономным образом в клетках CAR/LepR + , подавляя их дифференцировку в линию остеобластов, чтобы поддерживать их HSC-поддерживающая функция (рис.1). Точные характеристики клеток остеолинии, ответственных за поддержку ГСК, представляют большой интерес, поскольку они могут можно использовать для прогнозирования того, могут ли терапевтические стратегии, регулирующие коммитацию остеобластов, также использоваться для усиления образование ниш ГСК из ненишевых клеток.
Рисунок 1. КлеткиCAR/LepR + представляют собой МСК, окружающие синусоиды и/или вблизи эндоста.Они экспрессируют высокие уровни хемокина Cxcl12, ключевого HSC. фактор ниши, который вместе с Scf регулирует содержание и удержание ГСК в нише костного мозга. остеогенный и Потенциал адипогенной дифференцировки клеток CAR/LepR + ингибируется экспрессией факторов транскрипции Ebf1/3 , гарантируя, что эти специализированные МСК остаются недифференцированными, и позволяя поддерживать полость костного мозга, контролируя экспрессию Cxcl12 и Scf как хорошо физически сохраняя нишу HSC.
Биомиметическая инженерия in vitro гемопоэтической ниши человека с функциональными свойствами
Микроокружение костного мозга (КМ) отвечает за поддержание активности гемопоэтических стволовых клеток (ГСК), что позволяет производить зрелые клетки крови на протяжении всей жизни (1, 2). Регуляция самообновления и дифференцировки ГСК достигается сложными клеточными (3), молекулярными (4, 5), структурными (6) и физическими (7, 8) сигналами, определяющими нишу ГСК (2, 9).Компоненты ниши HSC человека и то, как эти элементы взаимодействуют, чтобы модулировать судьбу стволовых клеток, остаются плохо изученными.
Поле затруднено из-за ограниченных возможностей доступа и использования информации от человеческих особей. Химерные животные модели (10) наиболее точно воспроизводят физиологию человека in vivo, но в этих условиях эта ниша остается недоступной для экспериментальных манипуляций и оптических наблюдений (11, 12). Кроме того, межвидовой химеризм как в гемопоэтических клетках, так и в их окружении затрудняет интерпретацию экспериментальных результатов.Разработка заменителей in vitro является многообещающей альтернативой с превосходной настраиваемостью и производительностью (13, 14). Предыдущие исследования описывали комбинацию различных стромальных и гемопоэтических предшественников с использованием различных культуральных субстратов (9⇓–11), что приводило к фенотипическому сохранению специфических фенотипов крови. Тем не менее, повторение структурной организации BM, включая основные межклеточные и клеточно-матриксные взаимодействия (15⇓⇓⇓–19), и связанное с этим функциональное сохранение HSCs (20) все еще неуловимы.
Потребность в передовых культуральных системах более высокой биологической сложности получает все большее признание (21) для изучения фундаментальной биологии стволовых клеток. Подобно «органогенезу в чашке», предложенному для сложных органов [например, легких (22), молочной железы (23), почек (24) и печени (25)], инженерия in vitro среды БМ человека (21, 26 ⇓–28), способные поддерживать ГСК (28, 29), позволят их изучать в условиях, свободных от ксеногенов.
Здесь мы сообщаем о системе in vitro, поддерживающей разработку и обслуживание аналога BM человека.Наш подход заключается в использовании пористых гидроксиапатитовых каркасов со структурными и композиционными особенностями кости (30), функционализированных мезенхимальными стромальными клетками человека (hMSC) и внеклеточным матриксом (ECM), депонированным во время их постепенного созревания в остеобластную линию. Культивирование hMSC проводят при прямом перфузионном потоке (31), обеспечивая эффективное снабжение питательными веществами/удалением отходов, имитируя интерстициальный поток и связанное с ним напряжение сдвига. Кровяной компартмент вводили в полученную трехмерную стромальную ткань путем перфузии клеток CD34 + , полученных из очищенной пуповинной крови человека (CB).Этот сконструированный органоид частично повторяет структурные и функциональные особенности человеческого BM в определенных и настраиваемых условиях.
Results
Трехмерные микроокружения могут быть созданы внутри системы перфузионного биореактора.
Создание трехмерных микроокружений было выполнено путем дифференцировки первичных hMSC, полученных из костного мозга, на керамических материалах в перфузионном биореакторе. Клетки сначала метили трансгеном VENUS (>93%) ( SI Приложение , рис.S1 A ) для облегчения последующего анализа. Чтобы добиться инженерии остеобластоподобной стромы, мы приняли протокол, ранее использовавшийся для создания костных трансплантатов (32) (рис. 1 A ). hMSC сначала культивировали в течение 1 недели в пролиферативной среде (PM) для увеличения числа клеток и обеспечения колонизации каркаса, а затем в течение 3 недель добавляли остеогенную среду (OM) для стимулирования дифференцировки клеток при одновременной стимуляции продукции внеклеточного матрикса (ECM) (33). Полученная ткань была определена как «инженерная ниша» (eN) (рис.1 A ), в то время как голая керамика (Ce) (рис. 1 A ), не содержащая hMSC, но загруженная клетками CD34 + , использовалась в качестве внутреннего контроля 3D-культуры. Кровяной отсек впоследствии вводили путем инъекции клеток CD34 + человека, полученных из CB, от отдельных доноров в трубку устройства. Совместное культивирование in vitro поддерживали в течение 1 недели в бессывороточной среде с добавлением низких концентраций гемопоэтических цитокинов [10 нг/мл тромбопоэтина (ТПО), фактора стволовых клеток (SCF) и лиганда тирозинкиназы 3, родственного Fms (Flt3- л)].После извлечения из камеры биореактора Ce не имел очевидных структур ECM (Ce, SI, Приложение , рис. S1 B ), в то время как eN демонстрировал черты инженерной ткани с толстыми гелеобразными структурами, гомогенно покрывающими исходный материал. (eN, рис. 1 B ). Сканирующая электронная микроскопия подтвердила отложение внеклеточного матрикса, в котором заключены клетки, предположительно как стромального (фибробластическая форма, рис. 1 B ), так и кровяного происхождения (круглая форма, рис. 1 B ), включая делящиеся клетки (рис.1 В ).
Рис. 1.В системе перфузионного биореактора можно создавать трехмерные микроокружения. ( A ) Экспериментальный дизайн для создания трехмерных ниш в перфузионном биореакторе. ОМ — остеогенная среда; ПМ — пролиферативная среда; SFEM+GF, бессывороточная среда плюс факторы роста, фактор стволовых клеток, тромбопоэтин и лиганд Flt3. ( B ) Сконструированная ниша (eN, Left ) демонстрирует плотные гелеобразные структуры, гомогенно покрывающие исходный материал (Ce).Сканирующая электронная микроскопия изображений eN ( Right ) подтвердила отложение внеклеточного матрикса, который включает клетки, предположительно как стромального, так и кровяного происхождения. Стрелки указывают на наличие делящихся клеток.
Созданные трехмерные микроокружения позволяют поддерживать гемопоэтические стволовые клетки и клетки-предшественники с функциональными свойствами.
Чтобы охарактеризовать их клеточный состав, 3D-микросреда была обработана для извлечения клеток (эффективность извлечения> 92%, с общей гибелью клеток ниже 1.5%) ( SI Приложение , рис. S1 B и C ) и последующий количественный фенотипический анализ ( SI Приложение , рис. S1 D и E ).
eN состоял из более чем 4,7 × 10 6 клеток крови ( SI Приложение , рис. S1 F ) на биореакторную систему [61-кратное увеличение (fi) по сравнению с исходными 70 000 CD34 + клеток] ( Приложение SI , рис. S1 G ). Напротив, в отсутствие инженерной стромы размножение клеток крови было ограничено до 7.8 ф.и. ( SI Приложение , рис. S1 F и G ). Было показано, что при соотношении HSPCs над коммитированными клетками (CD34 — /CD38 + ) ниже, чем Ce (38 против 204) ( SI Приложение , рис. S1 H ), eN, как было показано, поддерживает распространение дифференцированных популяций. ЭН также способствовал поддержанию фенотипических гематопоэтических стволовых популяций и популяций предшественников (HSPCs) (рис. 2 A ), приводя к систематически более высокому общему количеству HSPCs (46.2 ф.и. против 6,4 для Ce), включая HSC (13,8 fi против 1,6), мультипотентные предшественники (MPP) (8,5 fi против 2,3) и мультипотентные лимфоидные предшественники (MLP) (161 fi против 20) ( SI Приложение , Рис. S2 A ).
Рис. 2.Спроектированные трехмерные микроокружения позволяют поддерживать HSPC с функциональными свойствами. ( A ) Сконструированная ниша (eN) поддерживает экспансию фенотипических гемопоэтических стволовых клеток и клеток-предшественников (HSPC), гемопоэтических стволовых клеток (HSC), мультипотентных клеток-предшественников (MPP) и мультипотентных лимфоидных клеток-предшественников (MLP), согласно количественной оценке анализ проточной цитометрии после 3D-культуры. n ≥ 8 биологических повторов. Ce, только керамика. *** Р < 0,001, **** Р < 0,0001. ( B ) Улучшенное поддержание колониеобразующего потенциала eN по сравнению с клетками CD34 + , культивируемыми Ce. BFU-E, взрывообразующая единица-эритроид; GEmM, колониеобразующая единица-гранулоцит, эритроид, макрофаг, мегакариоцит; ГМ, колониеобразующие единицы-гранулоциты и макрофаги. n ≥ 9 биологических повторов. ( C ) Долгосрочная репопуляционная способность eN и Ce культивируемых клеток CD34 + .Восстановление компартмента крови человека (процент клеток CD45 человека + в мононуклеарных клетках) у мышей NSG показано с помощью анализа периферической крови проточной цитометрией. Положительным контролем служили некультивируемые клетки CD34 + . n ≥ 4 биологических повторов. Клетки CD34 + человека, культивированные на eN и Ce, также прочно прижились в костном мозге и селезенке ( D ) трансплантированных мышей, что оценивалось с помощью проточной цитометрии через 28 недель после трансплантации. n ≥ 4 биологических повторов.
Способность hMSC поддерживать пролиферацию клеток CD34 + была дополнительно подтверждена в 2D-установке ( SI Приложение , рис. S2 B ). Используя недифференцированные hMSC в качестве фидерного слоя (2D hMSC) по сравнению с контролем без hMSC (2D), мы наблюдали значительное увеличение фенотипических HSPC (100 fi против 15), HSC (29 fi против 4), MPP (43 fi против 16) и MLP (279 fi против 21) ( SI Приложение , рис. S2 B ), в соответствии с результатами, полученными на хорошо зарекомендовавших себя культурах Dexter (34, 35).Способность образованной стромы сохранять функциональность культивируемых клеток крови оценивали как с помощью анализов in vitro, так и in vivo. Соответствующие донорские клетки до культивирования in vitro («некультивируемые») использовали в качестве функционального положительного контроля. Анализы колониеобразующих единиц in vitro продемонстрировали сохранение способности стволовых клеток или клеток-предшественников (с многолинейностью и потенциалом пролиферации) культивируемых клеток CD34 + , на что указывает образование миелоидных колоний с той же морфологией, что и их некультивируемые аналоги (приложение SI). , рис.S2 С ). Количество колоний было увеличено для клеток, извлеченных из 3D-культур, для колониеобразующих единиц-гранулоцитов и макрофагов (GM) (42,8 fi против 4,8 для Ce), взрывообразующих единиц-эритроидов (BFU-E) (113 fi против 18). , и колониеобразующая единица-гранулоциты (GEmM) (36,5 мкм против 4,8) колоний (рис. 2 B ).
Стволовой и прогениторный потенциал культивируемых клеток дополнительно тестировали путем внутрибедренной трансплантации равного количества клеток CD34 + из Ce или eN облученным мышам NSG.Кровяной компартмент человека был успешно восстановлен во всех группах [>1% CD45-положительных клеток человека (hCD45) в периферической крови] (Фиг. 2 C ). Химеризм был обнаружен уже на 6-й неделе и сохранялся в течение 28 недель после трансплантации, что указывает на долгосрочную способность трансплантированных клеток к репопуляции. Как и ожидалось, самый высокий химеризм был получен при трансплантации некультивируемых CD34 + , служащих положительным контролем (27,9 ± 3,1%) (фиг. 2 C ). Клетки CD34 + , полученные из Ce и eN, демонстрировали аналогичные уровни восстановления, в среднем 2.7 ± 0,8% и 6,4 ± 1% hCD45 соответственно, обнаруженные в периферической крови (рис. 2 C ). Химеризм также оценивали в БМ и селезенке животных с прочно привитыми клетками человека во всех условиях, с тенденцией к более высокой частоте hCD45, обнаруженной в eN, чем в группе Се, как в БМ (6,4% ± 0,5 против 5,2 % ± 2,5, рис. 2 D ) и селезенки (12,6 % ± 4,4 против 5,6 % ± 2,8, рис. 2 D ). Функциональность клеток CD34 + дополнительно оценивали по способности к реконституции мультиклонов.На 18-й неделе после трансплантации клетки из всех состояний успешно восстанавливали миелоидные и лимфоидные линии ( SI, Приложение , рис. S2 D ).
Эти данные показывают, что, по сравнению с аналогами Ce, eN дает гемопоэтические клетки с аналогичным потенциалом восстановления, но в большем количестве. Это говорит о том, что генерируемая ткань обеспечивает сигналы, способные усиливать поддержание/экспансию CFU-HSPC с потенциалом приживления трансплантата in vivo и восстановления нескольких линий.
Молекулярная характеристика спроектированной ниши показывает создание остеобластоподобной трехмерной среды.
Для выявления факторов, связанных с устойчивым развитием кроветворения, была дополнительно охарактеризована eN. Во-первых, мы отслеживали секрецию ключевых цитокинов на протяжении всего времени культивирования. Воспалительные факторы [интерлейкин 6 (IL-6), интерлейкин 8 (IL-8), макрофагальный колониестимулирующий фактор (MCSF) (рис. 3 A ) и моноцитарный хемоаттрактантный белок-1 (MCP-1)] показали самые высокие различия и были обнаружены при высоких концентрациях в eN.В частности, продукция IL-6 и IL-8 существенно увеличивалась при добавлении HSPC (eN, фиг. 3, —). Белки Flt3-L, TPO и SCF были обнаружены в концентрациях, сходных с концентрациями, добавленными в среду совместного культивирования ( SI, Приложение , рис. S3 A ), что позволяет предположить, что hMSC секретируют низкие уровни этих поддерживающих HSPC факторов. Фактор роста эндотелия сосудов α (VEGFα) и ангиопоэтин 1 (Ang-1) также были обнаружены на значительных уровнях (eN, SI, Приложение , рис.S3 A ), хотя Ang-1 со временем снижался и оставался стабильным после загрузки HSPC.
Рис. 3.Молекулярная характеристика спроектированной ниши показывает создание остеобластоподобной трехмерной среды. ( A ) В условиях искусственной ниши (eN) наблюдается более высокая концентрация воспалительных цитокинов, чем в керамическом состоянии (Ce), согласно анализу Luminex супернатантов 3D-культуры. ИЛ-6, интерлейкин 6; ИЛ-8, интерлейкин 8; MCP-1, моноцитарный хемотаксический белок 1; MCSF, колониестимулирующий фактор макрофагов. n ≥ 3 биологических повторов. Добавление клеток CD34 + в ниши индуцирует секрецию цитокинов только в условиях eN. ( B ) hMSCs несут основную ответственность за высокие уровни воспалительных цитокинов, обнаруженных в eN. Это было оценено с помощью анализа экспрессии генов клеток-предшественников крови (CD34 + ) и hMSC до (28-й день) и после совместного культивирования с клетками крови (35-й день). n ≥ 3 биологических повторов. ( C ) hMSCs приобретают генетический профиль остеобластоподобной ниши в культуре.Анализ экспрессии генов hMSC, полученных из eN (eN hMSC). hMSC (hMSC день 0) указывают основные уровни экспрессии генов до их 3D-культуры. ALP, щелочная фосфатаза; BSP, костный сиалопротеин; HIF1α, индуцируемый гипоксией фактор 1α. n ≥ 3 биологических повторов.
Чтобы получить более полное представление о клеточных компартментах, связанных с секрецией фактора, мы изолировали как кровяные предшественники (CD34 + ), так и мезенхимальные популяции до (определяется как hMSC на 28 день) ( SI Приложение , рис.S3 B ) и после кокультуры CD34 + (определяется как eN-hMSC 35 день, Приложение SI , рис. S3 B ). Это подтвердило сильную экспрессию воспалительных цитокинов (IL-6, MCSF) hMSC во время совместного культивирования с HSPC с уровнями, заметно более высокими, чем в клетках-предшественниках крови (Fig. 3 B ). IL-8 и MCP-1 экспрессировались как клетками крови, так и мезенхимальными клетками (фиг. 3 B ). Интересно, что после совместного культивирования с клетками CD34 + hMSC значительно увеличили экспрессию IL-6 и IL-8 (фиг.3 В ).
Чтобы оценить роль IL-6 и IL-8 в системе, мы исследовали эффект их добавления в условиях Ce ( SI Приложение , рис. S4 A ). IL-6 и IL-8 в дозах, соответствующих измеренным в eN, приводили к значительному увеличению числа HSPC, коммитированных предшественников (CD34 + /CD38 + ), гранулоцитарно-моноцитарных предшественников (GMP) и MLP ( Приложение SI , рис. S4 B ). Однако никакого влияния на HSC и MPP не наблюдалось ( SI, Приложение , рис.С4 Б ). Это предполагает, что эти воспалительные цитокины опосредуют пролиферацию коммитированных популяций в eN, тогда как наблюдаемое расширение компартмента стволовых клеток управляется другими факторами hMSC.
До загрузки CD34 + hMSC в сконструированной ткани преимущественно состояли не только из остеобластоподобных клеток, но также из пула, экспрессирующего маркеры-предшественники ( SI Приложение , рис. S5). Далее мы проанализировали профиль транскрипции eN-hMSC как нишевых клеток в конце культивирования (eN-hMSC, день 35) и сравнили его с hMSC после экспансии (hMSC, день 0) и hMSC в eN до загрузки CD34 + (hMSC день 28).Это подтвердило остеобластический профиль hMSC (35-й день) в конце 3D-культивирования, о чем свидетельствует активация щелочной фосфатазы (ALP) и костного сиалопротеина (BSP) (17 и 869 мкФ соответственно) (рис. 3 ). С ). Интересно, что заметное увеличение экспрессии Nestin (117 мкФ) было получено hMSC после совместного культивирования с клетками крови (рис. 3 C ), а также снижение индуцируемого гипоксией фактора 1-альфа (HIF1α, 8,9-кратное) выражение.
Таким образом, комбинируя анализ экспрессии белка и гена, мы описываем остеобластную (36, 37) и связанную с нишей (1) молекулярную подпись чМСК, связанную с провоспалительными свойствами, которая приобретается после кокультуры с кровяным компартментом.Это указывает на создание остеобластоподобной нишевой среды, взаимодействующей с гемопоэтическими клетками и способной регулировать активность HSPC, включая пролиферацию и функциональную регуляцию.
Спроектированная ниша имеет общие структурные и композиционные особенности с родным человеческим БМ.
Роль hMSC в создании благоприятной среды для гемопоэтического компартмента была дополнительно исследована путем изучения организации и состава eN.Иммунофлуоресцентный анализ срезов толстой конструкции выявил однородную сеть, образованную hMSCs внутри каркаса (сигнал VENUS, рис. 4 A ). Эта наблюдаемая мезенхимальная фракция состояла из 1,3 × 10 6 клеток, происходящих из hMSC (рис. 4 B ), по оценке проточной цитометрии. Генерируемая ткань, полученная в результате остеобластной дифференцировки hMSC, состояла из плотной стромы человека, заполняющей поры материала и включающей мезенхимальные клетки (рис. 4 C ).В состав депонированного ВКМ входят коллаген типа 1, коллаген типа 4 и фибронектин (рис. 4 C ), указанные как основные структурные белки костного мозга (38). Действительно, они также были в изобилии обнаружены в образцах BM, полученных от здоровых доноров (человеческая ниша, Fig. 4 D ). Сходство между человеческой тканью и созданной нами нишей не ограничивалось структурными белками. В биоптатах человека окрашивание остеокальцином выявило присутствие остеобластов, выстилающих поверхность кости (рис.4 Д ). Примечательно, что в eN аналогичная картина наблюдалась при обнаружении остеокальцина в клетках на границе между керамическим материалом и стромой клетки/ECM (рис. 4 C ).
Рис. 4.Сконструированная ниша имеет общие структурные и композиционные особенности с нативным БМ человека. ( A ) hMSCs образуют гомогенную клеточную сеть, распределенную внутри каркасного материала (конфокальная микроскопия, показаны репрезентативные изображения). ( B ) eN состоит из 1.3 × 106 клеток, происходящих из чМСК, в конце 3D-культуры (количественная оценка проточной цитометрией). n = 9 биологических повторов. Композиционное и структурное сходство внеклеточного матрикса в сконструированных нишах ( C ) и образцах костного мозга человека ( D ), оцененное с помощью конфокальной микроскопии. б — костная ткань (автофлуоресценция в синем канале). Стрелки указывают на присутствие остеокальцина в остеобластах, выстилающих поверхность кости.
В целом эти результаты свидетельствуют об успешном формировании сложной ткани при перфузионной культуре, связанной с развитием и поддержкой кроветворения человека (рис.2). Было показано, что сконструированная среда имеет общие структурные и композиционные особенности, типичные для нативного BM человека, что свидетельствует о частичном восстановлении нишевой среды, подобной остеобластам.
Созданная с помощью биореактора ниша показывает функциональную компартментализацию.
Разработанная культуральная система состоит из двух отдельных фаз: жидкой фракции, состоящей из супернатанта (SN) Приложение SI , рис. S6) и стромальной/ECM ткани, заключенной в камеру каркаса (ECM, Приложение SI , Инжир.С6). Это привело нас к дальнейшей гипотезе о том, что две среды могут по-разному влиять на распределение фенотипов клеток крови. Таким образом, мы отдельно проанализировали HSPC, полученные из этих двух фаз, с помощью проточной цитометрии ( SI, Приложение , рис. S6).
Несмотря на конвекцию, вызванную перфузионным потоком, наблюдалось специфическое распределение клеток, поскольку 80% извлеченных популяций HSPC (CD34 + /CD38 — ) находились в ВКМ (рис. 5 A , Left ).Напротив, более совершенные популяции (CD34 — / CD38 + ) демонстрировали сбалансированное распределение с 54% в ECM и 46% в SN (рис. 5 A , справа ). Отличительный паттерн может быть идентифицирован путем дальнейшей оценки предпочтительной локализации популяций стебля и предшественников в соответствии с их прогрессивной детерминацией (Fig. 5 B ). Примечательно, что HSC были обнаружены почти исключительно в строме (98%) (рис. 5 B ) вместе с подавляющим большинством MPP (76% в ECM) (рис.5 В ). Это относилось в меньшей степени к общим миелоидным предшественникам (CMP) и мегакариоцитарно-эритроидным предшественникам (MEP) (67% в ECM), в то время как GMP были более распространены в SN (61%) (рис. 5 B ). Также было показано, что локализация влияет на функциональность клеток in vitro, поскольку клетки CD34 + , извлеченные из внеклеточного матрикса, проявляли повышенную способность образовывать миелоидные колонии по сравнению с их аналогами SN (рис. 5 C ). Это включает превосходное формирование колоний GEmM, GM и BFU (2.6, 3.7 и 3.1 л.и. соответственно).
Рис. 5.Ниша, спроектированная биореактором, демонстрирует функциональную компартментализацию. ( A ) В биореакторной системе большинство HSPC локализовано в ткани (ECM), в то время как коммитированные клетки более равномерно распределены между ECM и супернатантом (SN). n ≥ 8 биологических повторов. ( B ) Популяции HSC, MPP и CMP/MEP преимущественно локализуются в строме (ECM), а не в жидкой фазе. n ≥ 8 биологических повторов.( C ) Потенциал формирования верхних миелоидных колоний клеток CD34 + , извлеченных из внеклеточного матрикса. BFU, взрывообразующая единица-эритроид; GEmM, колониеобразующая единица-гранулоцит, эритроид, макрофаг, мегакариоцит; ГМ, колониеобразующие единицы-гранулоциты и макрофаги. n ≥ 12 биологических повторов. ( D ) Конфокальный микроскопический анализ eN демонстрирует присутствие клеток CD34 + на разной глубине ткани ( справа , увеличение).Фракции CD34 + и hMSC были обнаружены внутри организованного внеклеточного матрикса ( E , слева ), в котором они установили физические взаимодействия ( E , справа ). Се, керамика; Col1, коллаген типа 1.
Присутствие клеток крови человека в нише было подтверждено с помощью иммунофлуоресценции, при этом человеческие клетки CD34 + в большом количестве обнаруживались в ЕСМ сконструированной ткани (рис. 5 D ). Клетки располагались преимущественно по периферии скаффолдов (край, рис.5 D ), но также в значительной степени инфильтрировали ткани, о чем свидетельствует присутствие во внутренних порах (внутренние, рис. 5 D ) и в более центральной области (центральные, рис. 5 D ). Важно отметить, что мезенхимальная и гемопоэтическая фракции в сконструированной строме были обнаружены внутри организованного ВКМ (рис. 5 E , слева ) и тесно взаимодействовали, о чем свидетельствуют явные физические контакты (рис. 5 E , справа ). ).
В целом это демонстрирует существование функционального разделения в трехмерной системе перфузии.eN демонстрирует свойство иерархического хемоаттрактанта в популяциях HSPC, в то время как детерминация постепенно ассоциируется с высвобождением в жидкую фазу. Преимущественное удержание стволовых клеток в строме вместе с доказательствами взаимодействий hMSC-CD34 + предполагает важную, запускаемую hMSC регуляцию активности HSPC, что приводит к превосходному сохранению функциональности стволовых клеток.
Ниши BM с индивидуальными молекулярными сигнатурами могут быть созданы с помощью генной инженерии.
Затем мы стремились доказать принцип использования нашей системы для создания индивидуальных гемопоэтических ниш с индивидуальной молекулярной сигнатурой.
С этой целью первичные hMSCs эффективно трансдуцировали с использованием лентивируса VENUS-SDF1α (>95%) ( SI Приложение , рис. S7 A и B ), что привело к сверхэкспрессии SDF1α ( SI Приложение , Рис. S7 C ). Затем были получены сконструированные SDF1α ниши (eN-SDF1α) в соответствии с ранее описанным протоколом и проведено сравнение с eN без обогащения SDF1α.На протяжении всего периода культивирования eN-SDF1α непрерывно продуцировал значительные уровни белка SDF1α (>2000 пг/мл/день/биореактор) (рис. 6 A ) по сравнению с eN (<300 пг/мл/день/биореактор). биореактор) (рис. 6 A ).
Рис. 6.Ниши костного мозга с индивидуальными молекулярными сигнатурами могут быть созданы с помощью генной инженерии. ( A ) Ниша, сконструированная с помощью SDF1α (eN-SDF1α, красные треугольники), секретирует более высокие уровни SDF1α, чем контрольная сконструированная ниша (eN, черные крестики) на протяжении всего периода 3D-культивирования. n ≥ 3 биологических повторов. ( B ) eN и eN-SDF1α оказались макроскопически идентичными ( слева ), но изображения конфокальной микроскопии выявили повышенное присутствие белка SDF1α в ткани eN-SDF1α. ( C ) Сравнение отношения абсолютного количества гемопоэтических популяций из eN и eN-SDF1α выявило идентичное содержание супернатанта (SN, , левый ), в то время как строма eN-SDF1α (ECM, , правый ) содержала уменьшенный количество HSPC, HSC и MPP. n ≥ 8. ( D ) Индивидуализация SDF1α значительно увеличивала процент покоящихся HSPC, HSCs и MPPs в ECM eN-SDF1α, что оценивалось анализом клеточного цикла соответствующих популяций. n = 5. ГСК – гемопоэтические стволовые клетки; HSPCs, гемопоэтические стволовые клетки и клетки-предшественники; MLP, мультилимфоидные предшественники; MPPs, мультипотентные предшественники. * Р < 0,05.
При извлечении из камер биореактора различные искусственные ткани были макроскопически идентичными (рис.6 В ). Иммунофлуоресцентный анализ выявил присутствие hMSC в обеих нишах, но eN-SDF1α продемонстрировал повышенное присутствие белка SDF1α, подтверждая ранее наблюдаемый характер секреции и подтверждая молекулярную настройку ниши. Белка было особенно много внутриклеточно внутри hMSC, непосредственно выстилающих поверхность керамики (eN-SDF1α, рис. 6 B и SI, Приложение , рис. S7 D ).
Количественный фенотипический анализ методом проточной цитометрии был проведен для оценки эффекта обогащения SDF1α в системе.Общее количество клеток крови, собранных из обоих компартментов (ECM и SN), было одинаковым в группах eN и eN-SDF1α ( SI, Приложение , рис. S8 A ). Однако заметное снижение количества HSPC наблюдалось в eN-SDF1α, что отражалось в значительном кратном увеличении HSC (1,9 fi) и MPP (1,4 fi) в системах eN по сравнению с eN-SDF1α. Важно отметить, что эффект SDF1α, по-видимому, ограничивается этими популяциями, поскольку MLP и общее содержание крови были одинаковыми в обеих нишах ( SI Приложение , рис.S8 А ). Интересно, что наблюдаемые различия коррелировали с компартментализацией системы. Действительно, клеточное содержание как eN, так и eN-SDF1α в SN было идентичным по составу (SN, Fig. 6 C ). Вместо этого в компартменте ECM eN было обнаружено большее количество HSPC, включая более высокое содержание в HSC и MPP (2,1 и 1,6 fi соответственно), в то время как, опять же, MLP и глобальные показатели крови не были затронуты.
Мы исследовали статус клеточного цикла популяций HSPC, HSC и MPP, извлеченных из наших трехмерных условий.Это выявило эффект индукции покоя SDF1α в eN-SDF1α со значительным увеличением HSPC, MPP и HSC в фазе G0 (рис. 6 D и SI Приложение , рис. S8 B и C). ). В частности, в клетках, извлеченных из ECM, мы измерили увеличение на 29,6%, 36,1% и 26,9% количества покоящихся HSPC, HSC и MPP соответственно (рис. 6 D ). Таким образом, наблюдаемые более низкие доли HSC и MPP в ECM ниш eN-SDF1α были обусловлены индукцией покоя сверхэкспрессией SDF1α.
Эти выводы иллюстрируют возможность использования описанной модели для разработки настраиваемых ниш BM, способных конкретно влиять на распределение и поведение HSC и MPP в системе.
Возмущение поведения HSC при моделировании травмы в спроектированной нише.
Далее мы оценили, может ли распределение клеток крови быть нарушено путем имитации повреждения ниши ( SI Приложение , рис. S9 A ). Перед загрузкой CD34 + eN подвергали воздействию блеомицина (eN-Bleo) в качестве сильнодействующего препарата, вызывающего повреждение ДНК (39).Препарат изменил метаболизм чМСК (снижение активности на 30%) ( SI Приложение , рис. S9 B ), но не их жизнеспособность в системе ( SI Приложение , рис. S9 C ).
В то время как обе ниши демонстрировали одинаковое содержание HSPC, общее количество HSC было значительно выше в eN-Bleo ( SI Приложение , рис. S9 D и E ). Наблюдаемое увеличение было результатом более высокого содержания HSC в ECM eN-Bleo (рис. 7 A ), в то время как состав крови был идентичен в SN как eN, так и eN-Bleo ( SI Приложение , рис.S7 А ). Клетки, выделенные из соответствующих ниш ECM, показали существенное снижение процента покоящихся HSCs в eN-Bleo (8% против 20% в eN) (Fig. 7 B ). Интересно, что это было ограничено HSCs, и клеточный цикл популяций HSPC и MPP не был затронут.
Рис. 7.Возмущение поведения ГСК при моделировании повреждения в инженерной нише. ( A ) Сконструированные ниши, подвергшиеся воздействию блеомицина (eN-Bleo), показали большее количество HSC и MPP в ECM.Напротив, повреждение блеомицином не влияло на состав крови SN. n ≥ 3. ( B ) Поврежденные ниши (eN-Bleo) продемонстрировали нарушенную способность поддерживать HSC в состоянии покоя, что оценивалось с помощью анализа клеточного цикла. n ≥ 3. ГСК, гемопоэтические стволовые клетки; HSPCs, гемопоэтические стволовые клетки и клетки-предшественники; MLP, мультилимфоидные предшественники; MPPs, мультипотентные предшественники. ** Р < 0,01.
Эти данные показывают, что воздействие блеомицина ухудшает способность hMSC поддерживать HSC в состоянии покоя, что приводит к их повышенной пролиферации.Таким образом, мы подтверждаем возможность использования нашей системы для изучения кроветворения человека в определенных сценариях, например, после травмы.
Обсуждение
Мы сообщаем об успешной инженерии in vitro BM-подобных тканей в системе перфузионного биореактора. Созданные ниши демонстрируют высокую биологическую сложность, захватывая структурные, композиционные и организационные особенности нативной остеобластической среды человека, что приводит к поддержке функций HSPC. Кроме того, с помощью проверенной молекулярной настройки 3D-ниши и разработки конкретных сценариев травм система была проверена как инженерная платформа BM с настраиваемыми свойствами.
Тканевая инженерия открывает новые возможности для исследования стволовых клеток, позволяя нам решать фундаментальные биологические вопросы, которые невозможно исследовать с помощью традиционных культуральных планшетов. Тем не менее, его применение для создания жизнеспособных сред BM in vitro остается сложной задачей из-за ограничений моделирования, связанных со сложностью ткани. Это включает в себя точно определенную пространственную организацию, клеточное разнообразие и комбинированную пролиферацию и поддержание функциональности кровяного компартмента.Поскольку существующие модели (14, 15) не повторяют все эти особенности без исключения использования животных (40), мы в качестве альтернативы предложили дизайн «органоподобной» ткани для поддержки развития и поддержания кроветворения.
Наша система предлагает ключевые преимущества по сравнению с существующими подходами. Во-первых, в отличие от синтетических материалов (41⇓-43), внеклеточный матрикс более точно воспроизводит нативную микросреду. Несмотря на существенные достижения в этой области, искусственные матрицы не могут воспроизвести распределение и разнообразие сигналов, существующих в естественном ВКМ, и не предлагают их подходящего и физиологического представления (44⇓–46).Более того, посредством генетических модификаций hMSC и их индивидуального профиля секретируемых факторов мы ввели понятие модульности, ранее достигнутое с помощью синтетических матриц (41⇓-43). Биологическая доставка определенных цитокинов клетками представляет собой непрерывный процесс, в отличие от экзогенного добавления в культуральную среду, потенциально связанного с проблемой стабильности во времени. Эта стратегия весьма актуальна при распространении на предполагаемые нишевые факторы для идентификации ключевых клеточных подмножеств/молекул, влияющих на поведение стволовых клеток (47).В этом отношении наличие разделения в нашей системе может быть использовано для решения конкретных вопросов. Они варьируются от возможности изучения хемоаттрактантных эффектов интересующих факторов до исследования механизмов, управляющих высвобождением стволовых клеток за пределы их ниши, и связанных с ними функциональных различий.
Несмотря на то, что наш подход основан на биологическом опыте, он не полностью отражает сложность своего аналога in vivo (9). Важными отсутствующими компонентами являются, например, сосудистые и нейронные сети, которые, как известно, являются регуляторами активности HSC (48⇓-50).Это требует изучения их интеграции в систему, хотя и требует создания условий культивирования, поддерживающих жизнеспособность нескольких типов клеток (51). Тем не менее, описанная модель достигла следующего шага в сложности, что иллюстрируется наблюдаемой самоорганизацией мезенхимальных и кровяных компартментов, что предполагает развитие прокси BM с органоидными чертами.
В отличие от микрожидкостных подходов, мы нацелились на разработку макромасштабных ниш, чтобы сочетать разумную пропускную способность и множественные показания.В то время как микрожидкостные платформы состоят из миниатюрных систем с высоким уровнем распараллеливания, ограниченный объем и количество ячеек сдерживают мультиплексный анализ. Вместо этого размер нашей системы был совместим с несколькими одновременными оценками, включая экспрессию генов, проточную цитометрию, визуализацию и необходимое определение in vivo функциональности стволовых клеток и клеток-предшественников.
Мы наблюдали в нашей системе значительное увеличение количества фенотипических HSC. Это было достигнуто в строгих условиях без ксеногенов, включая низкую концентрацию гемопоэтических цитокинов, и было подтверждено с использованием одного донора CB и нескольких первичных материалов hMSC.Тем не менее, функциональность HSC все еще нуждается в тщательной оценке с помощью анализа вторичной трансплантации и лимитирующих разведений, прежде чем заявлять о функциональной экспансии HSC. С этой целью разработка и оптимизация 3D-параметров могут быть приняты для проверки суспензионных культур золотого стандарта (52), которые в настоящее время требуют высоких уровней цитокинов (53, 54) и добавок агонистов (52). Некоторые из параметров, которые можно настраивать в культуральной платформе с трехмерной инженерией, включают типы нишевых клеток, их конкретную стадию дифференцировки, каркасный материал (например,например, структура/состав) или физические факторы, определяемые перфузионной системой (например, напряжение сдвига, уровни кислорода/гипоксии). Все они действуют согласованно с синергетическими и конкурирующими эффектами и могут быть использованы для модуляции судьбы стволовых клеток (55).
Наконец, наша система может быть адаптирована для воспроизведения патологических состояний (56). Использование hMSC и/или HSPC, полученных от пациентов, страдающих заболеваниями крови (например, лейкемией), может дать возможность смоделировать заболевание in vitro в полностью человеческих и идеально персонализированных условиях.Это может представлять собой мощный инструмент с широким спектром применений, от выявления факторов, нарушающих регуляцию ниши или функций крови, до скрининга лекарств для прогнозирования реакции конкретного пациента на определенные виды лечения.
Перспективное выделение некроветворных клеток ниши и их дифференциальные молекулярные взаимодействия с ГСК | Кровь
Чтобы изучить это явление более подробно, мы сосредоточились на эндотелиальных клетках и МСК, выбрав взаимодействия, которые включают лиганды с особенно высокой экспрессией в 1 из этих 2 типов нишевых клеток (рис. 6).Среди этих предсказанных клеточно-специфических взаимодействий верхней ниши мы обнаружили несколько ранее описанных, таких как CXCR4/CXCL12 (CXCR4/SDF1) и KIT/KITL (KIT/SCF), связанные с MSC, или NOTCh2/JAG1, CD44/Selectin E или Селектин L/селектин Е, связанный с эндотелиальными клетками. Кроме того, 2 нишевые клетки экспрессировали различные компоненты внеклеточного матрикса, которые могут взаимодействовать с одной и той же молекулой клеточной адгезии на HSPC, такие как коллаген (эндотелиальные клетки) и ламинин β1 (МСК), взаимодействующие с интегрином β1 (HSPC).Наш анализ также был информативен в отношении взаимодействий рецептор-лиганд, которые, как было показано, участвуют в гомеостазе HSPC, но для которых не был определен клеточный источник лиганда (например, Axl / Gas6 47 с GAS6, экспрессируемым с помощью МСК). Наконец, наши результаты указывают на роль антагонистов лигандов в регуляции HSPC. МСК экспрессируют высокие уровни как CXCL12, так и его антагониста CXCL14. Более того, наряду с экспрессией BMP4, BMP5 и BMP6, МСК также экспрессировали фоллистатин-1 ( Fstl-1 ), антагонист надсемейства трансформирующего фактора роста β, известный своим ингибированием лигандов BMP.Эти специфические для МСК лиганды были особенно сильно экспрессированы МСК, выделенными из костного мозга (дополнительная фигура 6), однако МСК костного мозга и кости также демонстрировали отчетливую экспрессию рецепторов (дополнительная таблица 4). В эндотелиальных клетках мы обнаружили высокую и специфическую экспрессию как Jag1 , лиганда Notch, так и Egfl7 , антагониста Notch3 . Таким образом, наш глобальный анализ молекулярных взаимодействий, специфичных для клеток-ниш, с HSPCs идентифицирует предполагаемые новые пары рецептор-лиганд между клетками-нишами и HSPC.Это также указывает на сложную сеть, в которой как положительные, так и отрицательные лигандные стимулы сбалансированы для поддержания функции HSC.
Ниша
Мы и другие показали, что провоспалительные цитокины, такие как IFNa, могут приводить к эффективной активации покоящихся HSC in vivo (Essers et al. , Nature , 2009). Напротив, в условиях культивирования in vitro IFNa оказывает ингибирующее действие на культивируемые HSC, приводя к блокированию пролиферации HSC даже в присутствии поддерживающего фидерного слоя.Эти данные указывают на роль ниши BM в индуцированной воспалением стимуляции HSC. Однако лежащий в основе этого молекулярный механизм плохо изучен. Поэтому мы сосредоточимся на выяснении роли ниши BM в активации HSCs, вызванной воспалительным стрессом. В BM HSCs окружены внеклеточным матриксом (ECM), соединяющим HSCs с другими структурами ниши, таким образом, являясь интересным кандидатом, облегчающим коммуникацию между HSCs и компонентами ниши при стрессе. Матрилины (Matns) составляют семейство адапторных белков ВКМ, которые соединяют различные компоненты ВКМ, образуя надмолекулярную структуру.Один из Matns, Matn4, сильно экспрессируется в HSC, и экспрессия антикоррелирует с увеличением пролиферативной активности от HSC к предшественникам. Анализ мышей, лишенных Matn4, не показал явных гематологических аномалий, таким образом, Matn4 не требуется для поддержания HSC и покоя в условиях гомеостаза.