Особенности ип: Индивидуальный предприниматель — что нужно знать чтобы зарегистрировать ИП
Особенности сдачи отчетности в ПФ для ИП
В первую очередь необходимо проверить и заполнить реквизиты. Чтобы открыть карточку с реквизитами, нажмите на Ваше краткое наименование в левой части экрана и нажмите кнопку «Изменить».
Нажмите на кнопку «Данные о налогоплательщике».
На вкладке «Параметры отчетности» проверьте наличие в поле «Регистрационный номер в ПФ» Вашего регистрационного номера ИП, как работодателя.
На вкладке «Доп. сведения» проверьте наличие в поле «Регистрационный номер ИП в ПФ» регистрационного номера ИП, присвоенного Вам как самозанятому населению.
После проверки реквизитов можно переходить к формированию отчетности. Зайдите в раздел «ПФР» на вкладку «Персонифицированная отчетность».
Укажите период и нажмите на «Новый отчет».
Выберите «Расчеты страховых взносов и индивидуальные сведения».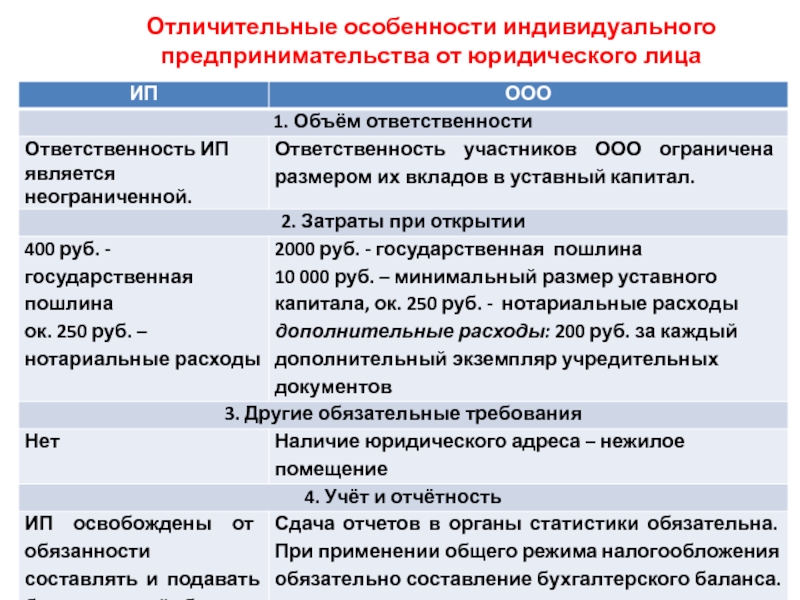
В открывшемся окне мастера в разделе «Выберите требуемую форму отчетности» нажмите на стрелку вниз для выбора требуемой формы.
Если вы формируете отчетность в качестве работодателя, то выберите «РСВ и индивидуальные сведения» и следуя по шагам мастера при помощи кнопки «Далее» сформируйте комплект отчетности.
Если вы формируете отчетность в качестве самозанятого населения (индивидуальные предприниматели, адвокаты, нотариусы и главы фермерских хозяйств) без найма к себе на работу других лиц, то выберите «Расчет страховых взносов» для того чтобы сформировать РСВ-2.
После выбора
После того как подготовите отчёт, нажмите «Передать на подпись» и отправьте его нажатием «Подписать и отправить».
ЭДО для ИП — Диадок
Зачем ЭДО для ИП
Электронный документооборот — это обмен электронными документами по телекоммуникационным каналам связи, которые обеспечивает оператор ЭДО. Такой формат делопроизводства оптимизирует бизнес-процессы, что нужно как закрепившимся на рынке компаниям, так и малому бизнесу. Ведь каждый ИП на любой системе налогообложения обязан оформлять хозяйственные операции документально, вести кадровый, налоговый, бухгалтерский учет.
Индивидуальным предпринимателям ЭДО с контрагентами поможет:
- создавать, получать, обрабатывать, отправлять, редактировать, согласовывать и подписывать документы;
- уменьшить административные расходы на бумагу, печать, обслуживание принтеров;
- сократить время на подготовку, доставку документов;
- отвечать за качество и полноту сведений в бухгалтерской отчетности;
- освободить время на отличные от заполнения бумаг задачи;
- вести кадровый учет, КУДИР (книгу учета доходов и расходов) и иные виды учета;
- контролировать работу сотрудников;
- корректно оформлять документы благодаря наличию в системе ЭДО стандартных форм.


Кроме того, онлайн можно регистрировать онлайн-кассы в ФНС и заключать договоры с оператором фискальных данных, а также участвовать в торгах на федеральных и коммерческих площадках, в проведении закупок по 44-ФЗ.
Электронный документооборот упростит и отправку отчетности в контролирующие госорганы — ФНС, Пенсионный фонд, Соцстрах, Росстат и другие. В электронном виде можно сдать декларацию по налогу на УСН, форму 4-ФСС. Только в электронном формате принимаются сведения о деятельности ИП в Росстат, справки 2-НДФЛ, если работников больше десяти, декларация по НДС.
Кроме того, обязательна безбумажная отчетность в ПФР и ФСС, если под вашим руководством работает более 25 человек, и в ФНС, если в компании числится 100 и более сотрудников. Остальным организациям электронная отчетность рекомендована.
Преимущества ЭДО для ИП
Преимущества ЭДО привлекают средний и малый бизнес. Благодаря электронному делопроизводству компании сокращают издержки, повышают общую эффективность всех процессов и становятся более конкурентоспособными на рынке.
Это происходит благодаря тому, что в отличие от бумажного делопроизводства ЭДО позволяет:
- Экономить деньги на печати, канцелярии, бумаге, забыть про почтовые, курьерские услуги.
- Редактировать документы, не допускать их дублирования, отслеживать их статус и сроки подписания.
- Забыть о трудностях, связанных с пересылкой оригиналов документов.
- Избегать ошибок за счет автопроверки форматов и сертификата электронной подписи, оперативно находить причину и исправлять расхождение данных.
- Хранить документы без затрат на содержание архива.
- Проводить сделки в кратчайшие сроки, быстро оформлять первичные, закрывающие документы.
- Без проблем предъявлять документы по требованию контролирующих органов.
- Обеспечить минимальный риск порчи, утери файлов.
- Увеличить скорость поступления данных в учетные системы.
- Повысить безопасность данных — передача происходит через защищенное соединение, а доступом к файлам владеет конкретное число пользователей.

Безбумажное делопроизводство повышает и производительность труда сотрудников. При бумажном документообороте работники тратят до половины рабочего времени на обработку документов. С ЭДО процессы ускоряются, а доступ к файлам легко получить даже удаленно.
Более того, систему ЭДО можно интегрировать с 1С и другими программами учета. Это позволит автоматически заполнять формы документов, настраивать проводку входящих файлов и дальнейшее формирование отчетности.
Откройте все возможности ЭДО для ИП с Диадоком
ПодключитьсяКакими документами ИП может обмениваться через ЭДО
Уже сегодня индивидуальные предприниматели переходят на формирование и сдачу налоговой и иной отчетности в электронном виде, а также на обмен следующими документами:
- договорами, соглашениями и приложениями к ним;
- счетами, счетами-фактурами, накладными, актами выполненных работ, УПД.

С помощью стандартных форм в редакторе Диадока можно создавать и заполнять счета-фактуры, накладные, акты, УПД, отправлять их, получать, изменять, согласовывать, подписывать, хранить. Отметим, что счета-фактуры будут равнозначны бумажным, только если составлены по утвержденному формату ФНС, переданы через оператора ЭДО и подписаны электронной подписью.
Как подключить ЭДО для ИП
Для организации электронного документооборота понадобится выбрать систему и оператора ЭДО, проанализировать делопроизводство, внести изменения в учетную политику и договориться об обмене электронными документами с контрагентами.
Способ организации ЭДО выбирается с учетом количества отправляемых документов и занятых в документообороте сотрудников:
- Веб-версия. Подойдет для малого документооборота: ее использование упростит переход на ЭДО, поскольку она работает в любом браузере и интерфейсом напоминает электронную почту.
- Модуль для учетной системы 1С. Интеграция системы ЭДО и 1С позволит работать в привычной системе.
- Интеграция с другими учетными системами.

Также при выборе системы ЭДО стоит обратить внимание на условия внедрения, набор опций, тарифы, наличие роуминга, режим работы техподдержки.
Поможем подобрать решение и подключиться к ЭДО
Оставить заявкуЗачем нужна электронная подпись для ЭДО
Электронная подпись (ЭП) в электронном документообороте необходима для того, чтобы подписанные документы имели ту же юридическую силу, что и их бумажные аналоги. ЭП визирует документ, подтверждает авторство подписи, гарантирует отсутствие изменений в документе после подписания.
В законе 63-ФЗ «Об электронной подписи» обозначены три вида ЭП:
- Квалифицированная (КЭП). Только эта подпись равнозначна собственноручной. Это самая защищенная ЭП: ее технология контролируется ФСБ, выдать ее может только аккредитованный Минцифры удостоверяющий центр (УЦ), а работает она только со средством криптографической защиты информации.
- Неквалифицированная (НЭП). В технологии тоже используются криптографические алгоритмы, а сама ЭП позволяет проверить личность подписанта и отсутствие в файле изменений. Для получения такой подписи можно обратиться в любой УЦ, а уровень ее безопасности ниже. Для обмена документами с контрагентами понадобится взаимное соглашение.
- Простая. Эта подпись подтверждает, что была сформирована конкретным человеком. Чаще всего применяется в онлайн-банкинге, интернет-магазинах, внутреннем документообороте. Простую подпись нельзя использовать для отправки документов в госорганы, для обмена с партнерами нужно соглашение.
 Только эта подпись равнозначна собственноручной. Это самая защищенная ЭП: ее технология контролируется ФСБ, выдать ее может только аккредитованный Минцифры удостоверяющий центр (УЦ), а работает она только со средством криптографической защиты информации.
Только эта подпись равнозначна собственноручной. Это самая защищенная ЭП: ее технология контролируется ФСБ, выдать ее может только аккредитованный Минцифры удостоверяющий центр (УЦ), а работает она только со средством криптографической защиты информации.В Диадоке компании-резиденты РФ для обмена документами пользуются только квалифицированной ЭП.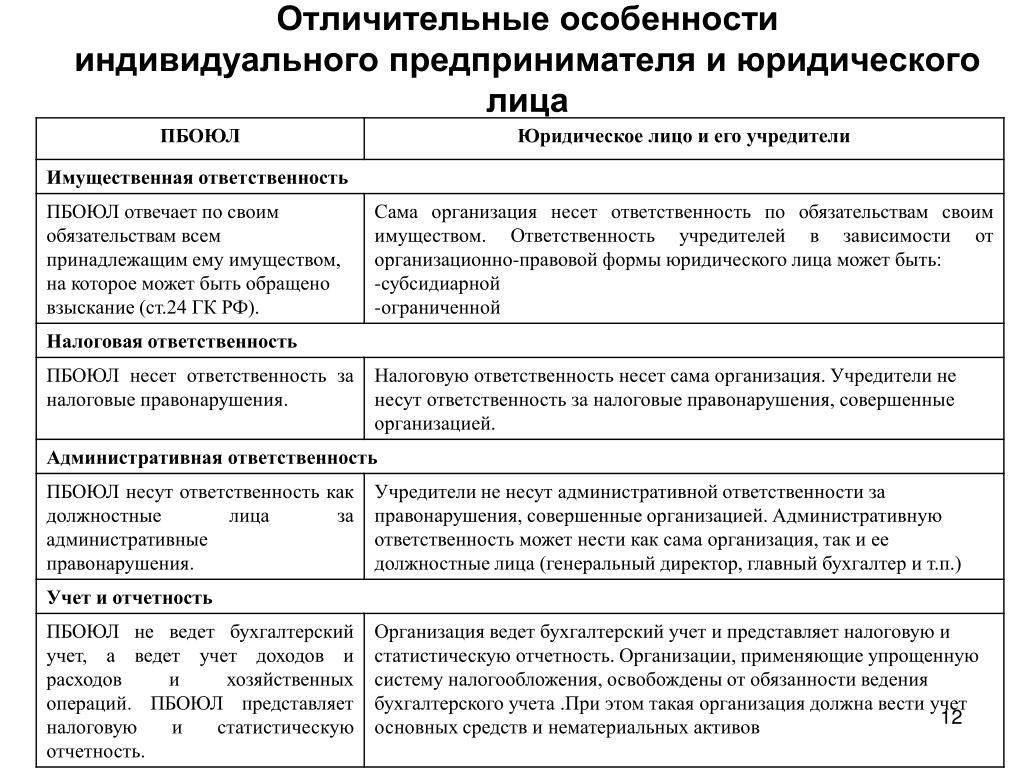 С ней для организации юридически значимого документооборота не понадобятся дополнительные соглашения с контрагентами, при этом подпись будет равнозначна собственноручной. Соглашение требуется только для организации международного ЭДО с контрагентом из другой страны, который использует НЭП.
С ней для организации юридически значимого документооборота не понадобятся дополнительные соглашения с контрагентами, при этом подпись будет равнозначна собственноручной. Соглашение требуется только для организации международного ЭДО с контрагентом из другой страны, который использует НЭП.
С КЭП предприниматель сможет вести документооборот, отчитываться онлайн и выполнять любые другие операции, которые нужно заверять личной подписью:
- Обмениваться с партнерами как формализованными документами (счета-фактуры, УПД, их корректировки), так и неформализованными (договоры, соглашения, счета).
- Обеспечить оперативное достижение договоренностей и проведение расчетов по сделкам.
- Отчитываться за год, квартал и другие периоды в контролирующие госорганы — ФНС, ПФР, ФСС, Росстат и т. д.
- Участвовать в электронных торгах, тендерах, госзакупках.
- Регистрировать онлайн-кассы.
При создании электронной подписи пользователь получит квалифицированный сертификат и два ключа — открытый для проверки подписи и закрытый для ее генерации.
Возможен ли обмен документами между двумя системами
Если у ИП и контрагента разные системы ЭДО, обмен документами тоже возможен. Для этого операторы должны поддерживать технологию роуминга. Документооборот при этом происходит так же, как и в рамках одной системы.
Настройка роуминга, как правило, не занимает много времени. Для подключения к нему в Диадоке нужно:
- На странице «Состояние роуминга между Диадоком и другими операторами» проверить, настроен ли уже роуминг с оператором интересующего контрагента.
- Заполнить заявку на подключение, дождаться выполнения.
С некоторыми операторами действует автоматическая настройка роуминга. В этом случае не понадобится даже заявка — пользователь может отправить контрагенту приглашение к обмену документами, как обычно. После принятия заявки…
Ответим на вопросы и настроим электронный документооборот
Подключиться|
Наименование параметра |
Значение |
Предел допускаемой приведенной основной погрешности, % |
±0,25 |
Частота входного сигнала синусоидальной формы, Гц |
от 45 до 1000 |
Напряжение питания постоянного тока, В: |
от 18 до 42 |
Выходной сигнал ( допустимое сопротивление нагрузки) |
0…10 В ( ³ 2 кОм)0…5 мА (0…2,5 кОм)0. .20 мА (0…1 кОм) .20 мА (0…1 кОм)
4..20 мА (0…1 кОм) |
Гальваническая изоляция входа – выход — питание |
есть |
Время установления выходного сигнала, с |
0,25 |
Средний срок службы не менее, л |
12 |
Средняя наработка на отказ, ч |
50000 |
Габариты, В×Ш×Г, мм |
75×22,5×108 |
Масса, не более, кг |
0,25 |
Температура эксплуатации,°С |
0…+60 для исполнения В4;-40…+60 для исполнения С4 |
Влажность |
до 80% при +35 °С и более низких температурах, без конденсации влаги |
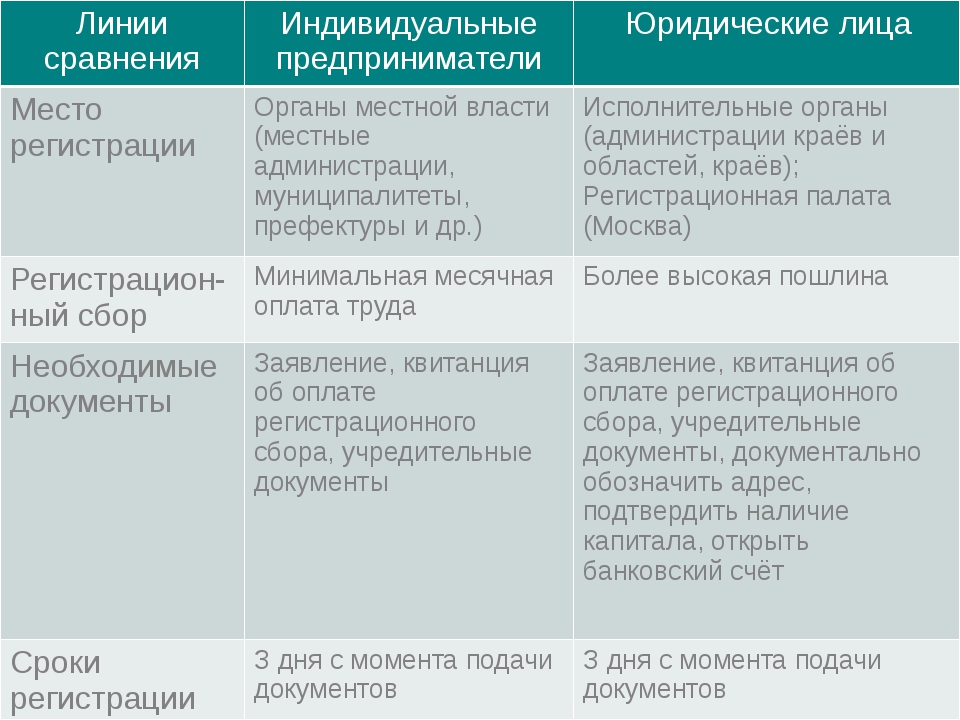
Порядок и особенности получения статуса индивидуального предпринимателя
По действующему законодательству, в целях извлечения прибыли, гражданин может создать юридическое лицо, или заниматься коммерческой деятельность без образования такового.
Стоит отметить, что статус индивидуального предпринимателя в правовом смысле неоднозначен, поскольку к нему одновременно применяются положения законодательства, распространяемое на физических лиц, а также на субъектов экономической деятельности.
Индивидуальными предпринимателями признаются зарегистрированные в установленном порядке физические лица, осуществляющие коммерческую деятельность без образования юридического лица. На территории России индивидуальным предпринимателем может быть как гражданин России, так иностранный гражданин, имеющий вид на жительство или разрешение на временное проживание на территории России.
В предыдущем законодательстве РФ использовались однородные понятия, такие как «частный предприниматель», «предприниматель без образования юридического лица» и т. д.
Главная особенность такого вида деятельности – гражданин отвечает по своим обязательствам собственным имуществом, что отличает его от участника общества с ограниченной ответственностью, где он отвечает по своим долгам и обязательствам в пределах внесенной доли в уставный капитал. Также, ИП не может получить некоторые виды лицензий (например, на розничную продажу алкоголя). Можно, сказать, что это единственный недостаток данной формы экономической деятельности. Но наряду с ним, статус индивидуального предпринимателя имеет множество достоинств. Например, простая процедура регистрации и ликвидации, легкий порядок принятия решений, упрощенный порядок предоставления отчетности в государственные органы, низкие штрафные санкции, относительно невысокие расходы.
Также, ИП не может получить некоторые виды лицензий (например, на розничную продажу алкоголя). Можно, сказать, что это единственный недостаток данной формы экономической деятельности. Но наряду с ним, статус индивидуального предпринимателя имеет множество достоинств. Например, простая процедура регистрации и ликвидации, легкий порядок принятия решений, упрощенный порядок предоставления отчетности в государственные органы, низкие штрафные санкции, относительно невысокие расходы.
Не стоит забывать и еще одну особенность, касаемую судебной защиты индивидуального предпринимателя. По общему правилу, защита прав ИП осуществляется в Арбитражных судах. Но если спор вытекает из гражданско-правовых отношений предпринимателя, как физического лица, то такие дела будут рассматриваться в судах общей юрисдикции.
Заниматься предпринимательской деятельностью могут только дееспособные совершеннолетние граждане. Статус ИП гражданин получает вследствие государственной регистрации.
Чтобы получить статус предпринимателя необходимо пройти процедуру государственной регистрации. Для этого требуется минимум действий. Необходимо предоставить определенный комплект документов в налоговый орган по месту регистрации гражданина. Пакет документов включает в себя: заявление формы Р21001, заверенное у нотариуса, заверенная у нотариуса копия паспорта, квитанция об уплате госпошлины. В заявлении помимо паспортных данных гражданина указываются коды видов экономической деятельности, которыми гражданин намерен заниматься. Документы в налоговый орган может подать гражданин лично, или обратиться в юридическую фирму, где это сделают за него. Также разрешается посылать документы по почте.
Срок государственной регистрации лица в качестве индивидуального предпринимателя составляет три рабочих дня, не считая дня подачи документов и дня выдачи.
На практике нередко встречаются случаи, когда налоговая отказывает в государственной регистрации. Причин тому может быть множество. Основными являются непредставление необходимого пакета документов, неверных или ложных сведений, гражданин был признан банкротом менее, чем год назад.
Причин тому может быть множество. Основными являются непредставление необходимого пакета документов, неверных или ложных сведений, гражданин был признан банкротом менее, чем год назад.
После государственной регистрации и получения всех необходимых документов, ИП имеет право приступить к осуществлению коммерческой деятельности, то есть заключает сделки, закупает товар, оказывает услуги.
Если Вы решили получить статус индивидуального предпринимателя и у Вас вызывает затруднение подготовка пакета документов, то лучше всего обратиться за помощью к квалифицированному специалисту, который все сделает за Вас.
Союз предпринимателей Южноприморский — Особенности учета при осуществлении нескольких видов деятельности ИП
Особенности заполнения декларации УСН
1. Если ИП осуществляет несколько видов экономической деятельности, то в титульном листе декларации по налогу, уплачиваемому в связи с применением УСН, проставляется код по ОКВЭД (ОК 029-2001) по виду деятельности, доходы от которой облагаются в рамках упрощенной системы налогообложения. (Минфин РФ письмо № 03-11-11/186 от 09.06.2012)
(Минфин РФ письмо № 03-11-11/186 от 09.06.2012)
2. Если ИП наряду с деятельностью, в отношении которой им применяется УСН, осуществляет иную предпринимательскую деятельность на основе патента (патентов), то он ведет налоговый учет по каждому основанию отдельно. (ФНС РФ письмо № ЕД-4-3/8971@ от 31.05.2012).
Учет доходов и расходов ИП ведет:
1. по деятельности, в отношении которой применяется УСН, в Книге учета доходов и расходов организаций и ИП, применяющих УСН (приложение № 1 к приказу Минфина России от 31.12.2008 № 154н).
2. по деятельности, осуществляемой на основе патента, в Книге учета доходов ИП, применяющих УСН на основе патента (приложение № 3 к приказу Минфина России от 31.12.2008 № 154н).
Раздельный учет по суммам уплачиваемых страховых взносов за работников
Уменьшить налог, уплачиваемый в связи с применением УСН, и стоимости патента на сумму страховых взносов можно при следующих условиях:
1. ИП должен вести раздельный учет по суммам уплачиваемых страховых взносов за работников, занятых в осуществлении деятельности, в отношении которой применяется УСН, и за работников, привлеченных по деятельности, осуществляемой на основе патента.
ИП должен вести раздельный учет по суммам уплачиваемых страховых взносов за работников, занятых в осуществлении деятельности, в отношении которой применяется УСН, и за работников, привлеченных по деятельности, осуществляемой на основе патента.
2. В случае невозможности разделения расходов на уплату страховых взносов (в том числе, за себя исходя из стоимости страхового года) между видами предпринимательской деятельности, распределение сумм страховых взносов производится пропорционально размеру доходов, полученных от соответствующих видов деятельности, в общем объеме доходов от осуществления всех видов предпринимательской деятельности.
3. Если ИП, не имеющий наемных работников, совмещает УСН и УСН на основе патента, общая сумма страховых взносов, уплаченных за себя исходя из стоимости страхового года, относится ко всем видам предпринимательской деятельности. (ФНС РФ письмо № ЕД-3-3/2476@ от 17.07.2012) Поэтому ИП вправе самостоятельно распределить фактически уплаченную стоимость страховых взносов на уменьшение налога, уплачиваемого в связи с применением УСН, и патента при оплате оставшейся части его стоимости.
ИП, не имеющий наемных работников, может
1. сначала уменьшить стоимость патента (после оплаты 1/3 стоимости патента, при оплате оставшейся ее части) на сумму страховых взносов, уплаченных до срока уплаты оставшейся части стоимости патента,
2. затем сумму налога по УСН, исчисленного за налоговый (отчетный) период, уменьшить на оставшуюся сумму страховых взносов, уплаченных (в пределах исчисленных сумм) за этот же период времени.
Некоторые особенности учета у ИП при сдаче имущества в аренду
ИП сдает в аренду оборудование белорусской организации, применяет общую систему налогообложения с уплатой НДС и ведением учета в книге учета доходов и расходов индивидуальных предпринимателей, согласно постановлению МНС от 24.12.2014 № 42 (далее – Книга). Выручка от реализации определяется по оплате. Стоимость обязательств (арендной платы) выражена в BYN эквивалентно определенной сумме USD. Согласно договору оплата производится до 20-го числа календарного месяца, следующего за отчетным, в BYN по курсу USD, установленному Нацбанком на дату оплаты.
Стоимость обязательств (арендной платы) выражена в BYN эквивалентно определенной сумме USD. Согласно договору оплата производится до 20-го числа календарного месяца, следующего за отчетным, в BYN по курсу USD, установленному Нацбанком на дату оплаты.
По состоянию на 31.12.2017 г. числится дебиторская задолженность по арендной плате за оборудование: сентябрь 2017 г. – 14 840 USD; октябрь 2017 г.– 24 000 USD; ноябрь 2017 г. – 24 000 USD; декабрь 2017 г. – 24 000 USD.
В январе 2018 г. поступила оплата:
– 09.01.2018 г. – 109 337,68 BYN (54 740 USD по курсу на 09.01.18 г. – 1,9974).
– 10.01.2018 г. – 34 578,88 BYN (17 200 USD по курсу на 10. 01.18 г. – 2,0104).
01.18 г. – 2,0104).
По задолженности за сентябрь 2017 г. выставлена ЭСЧФ по сроку на 60-й день с момента оказания услуг (29.11.2017 г.). По задолженности за октябрь 2017 г. выставлена ЭСЧФ по сроку на 60-й день с момента оказания услуг (03.01.2018 г.).
Должен ли ИП выраженную в BYN эквивалентно определенной сумме в USD сумму дебиторской задолженности, возникшей в 2017 г. и оставшейся непогашенной на конец года, пересчитать в BYN по курсу Нацбанка на 31 декабря 2017 г.?
Должен ли ИП пересчитывать согласно постановлению Минфина от 10.08.2017 № 23 (далее – постановление № 23) с 1 января 2018 г. выраженную в BYN эквивалентно определенной сумме в USD стоимость дебиторской и кредиторской задолженности на дату совершения операции…
Автор публикации:
Майя ЯРОШ, экономист- Статья
- 19 минут на чтение
Пожалуйста, оцените свой опыт
да Нет
Любая дополнительная обратная связь?
Отзыв будет отправлен в Microsoft: при нажатии кнопки отправки ваш отзыв будет использован для улучшения продуктов и услуг Microsoft.Политика конфиденциальности.
Представлять на рассмотрение
Спасибо.
В этой статье
ПРИМЕНЯЕТСЯ К: 2013 2016 2019 Выпуск по подписке SharePoint в Microsoft 365
Узнайте о новых функциях и обновлениях существующих функций в SharePoint Server 2016.
Для сравнения локальных функций SharePoint в выпусках SharePoint 2013 и SharePoint Server 2016 см. раздел Доступность функций SharePoint в локальных решениях.Новые функции SharePoint Server 2016 для конечных пользователей см. в статье Что нового в SharePoint Server 2016.
в статье Что нового в SharePoint Server 2016.
Обзор характеристик
В следующей таблице представлена сводка новых функций, которые вы можете опробовать в этом выпуске SharePoint Server 2016.
| Элемент | Описание | Подробнее |
|---|---|---|
| Услуги доступа | Новые функции Access доступны при развертывании служб Access в SharePoint Server 2016. | Дополнительные сведения см. в разделе Службы Access плюс клиент и сервер Access. |
| Характеристики соответствия | Новые функции соответствия для SharePoint Server 2016 включают политики удаления документов и хранения на месте. | Дополнительные сведения см. в разделе Функции соответствия. |
| Индивидуальные веб-части | Улучшено время компиляции настраиваемых файлов XSLT, используемых для запроса содержимого, сводных ссылок и веб-частей оглавления. | нет данных |
| Доступ к библиотеке документов | SharePoint Server 2016 включает новые функции специальных возможностей библиотеки документов. | Дополнительные сведения см. в разделе Специальные возможности библиотеки документов. |
| Прочные звенья | URL-адреса на основе ресурсов теперь сохраняют ссылки при переименовании или перемещении документов в SharePoint. | нет данных |
| Зашифрованные соединения | SharePoint Server 2016 поддерживает TLS 1.2 шифрование соединения по умолчанию. | Дополнительные сведения см. в разделе Зашифрованные соединения. |
| Быстрое создание коллекции веб-сайтов | Функция быстрого создания семейства веб-сайтов — это быстрый метод создания семейств веб-сайтов и сайтов в SharePoint. | Дополнительные сведения см. в разделе Быстрое создание семейства веб-сайтов. |
| Имена файлов — расширенная поддержка специальных символов | SharePoint Server 2016 теперь поддерживает использование некоторых специальных символов в именах файлов, которые были заблокированы в предыдущих версиях. | Дополнительные сведения см. в разделе Имена файлов — расширенная поддержка специальных символов. |
| Гибрид в SharePoint 2016 | Hybrid в SharePoint Server 2016 позволяет интегрировать локальную ферму с возможностями повышения производительности Microsoft 365, позволяя вам внедрять облако в своем собственном темпе. | Дополнительные сведения см. в разделе Гибрид в SharePoint Server 2016. |
| Идентификация и поиск конфиденциального контента | SharePoint Server 2016 теперь предоставляет те же возможности предотвращения потери данных, что и Office 365. | Дополнительные сведения см. в статье Определение и поиск конфиденциального содержимого в документах SharePoint Server 2016 и OneDrive. |
| Предварительный просмотр изображений и видео | Теперь вы можете просматривать изображения и видео в библиотеках документов SharePoint Server 2016. | Дополнительные сведения см. в разделе Предварительный просмотр изображений и видео. |
| Управление правами на информацию | SharePoint Server 2016 предоставляет возможности управления правами на доступ к данным (IRM) для защиты информации путем шифрования и защиты информации о библиотеках SharePoint с помощью OneDrive. | Дополнительные сведения см. в разделе Управление правами на доступ к данным. |
| Подставка для больших файлов | SharePoint Server 2016 теперь поддерживает отправку и скачивание файлов размером более 2047 МБ. | Дополнительные сведения см. в разделе Поддержка больших файлов. |
| Минимальная роль | MinRole — это новая функция в SharePoint Server 2016, которая позволяет администратору фермы SharePoint определять роль каждого сервера в топологии фермы. | Дополнительные сведения см. в разделе Топология фермы MinRole. |
| Мобильный опыт | SharePoint Server 2016 предлагает улучшенную мобильную навигацию. | Дополнительные сведения см. в разделе Мобильные возможности. |
| Новые функции в ПО от ноября 2016 г. для SharePoint Server 2016 (пакет функций 1) | Общедоступное обновление для SharePoint Server 2016 (пакет дополнительных компонентов 1) за ноябрь 2016 г. предлагает семь новых функций для SharePoint Server 2016. | Дополнительные сведения см. в статье Новые функции в PU от ноября 2016 г. для SharePoint Server 2016 (пакет дополнительных компонентов 1). для SharePoint Server 2016 (пакет дополнительных компонентов 1). |
| Новые элементы управления для работы с OneDrive | SharePoint Server 2016 предоставляет элементы управления в верхней части ваших личных папок документов, которые делают общие задачи в OneDrive более доступными. | Дополнительные сведения см. в статье Новые элементы управления для работы с OneDrive. |
| Новая корзина в OneDrive и на сайтах групп | SharePoint Server 2016 добавляет ссылку на корзину в левой области навигации на сайтах OneDrive и Team. | нет данных |
| Открытый формат документа (ODF) | В SharePoint Server 2016 добавлена поддержка файлов Open Document Format (ODF) для использования в шаблонах библиотеки документов. | Дополнительные сведения см. в разделе Формат открытого документа (ODF), доступный для библиотек документов. |
| Сервер проектов | Новые функции Project Server доступны в SharePoint Server 2016. | Дополнительные сведения см. в статье Project Server 2016. |
| Поддержка файловой системы ReFS | SharePoint Server 2016 теперь поддерживает диски, отформатированные в файловой системе ReFS. | Дополнительные сведения о файловой системе ReFS см. в разделах Обзор отказоустойчивой файловой системы и Устойчивая файловая система. |
| Бизнес-аналитика SharePoint | SharePoint Server 2016 теперь поддерживает SQL Server 2016 CTP 3.1, надстройку Power Pivot и Power View. | Дополнительные сведения о бизнес-аналитике SharePoint см. в статье Надстройка Power Pivot и Power View теперь доступны для использования с SharePoint Server 2016. |
| Поиск SharePoint | Приложение SharePoint Search Server претерпело значительные изменения в развертывании. | Дополнительные сведения см. в разделе Приложение службы поиска SharePoint. |
| Обмен улучшениями | В SharePoint Server 2016 доступно множество новых улучшений общего доступа. | Дополнительные сведения см. в разделе Общий доступ. |
| Просмотр папок сайта | SharePoint Server 2016 предоставляет новое представление «Папки сайта», которое позволяет вам получать доступ к библиотекам документов на сайтах, на которые вы подписаны. | Дополнительные сведения см. в разделе Представление папок сайта. |
| Закрепление страницы сайта | Эта новая функция помогает просматривать сайты и подписываться на них. | Дополнительные сведения см. в разделе Закрепление страницы Сайтов. |
| Шифрование соединения SMTP | SharePoint Server 2016 поддерживает отправку электронной почты на SMTP-серверы, использующие шифрование соединения STARTTLS . | Дополнительные сведения см. в разделе Шифрование SMTP-соединения. |
| Порты SMTP (не по умолчанию) | В SharePoint Server 2016 добавлена поддержка SMTP-серверов, которые используют порты TCP, отличные от порта по умолчанию (25). | Дополнительные сведения см. в разделе Использование портов SMTP, отличных от портов по умолчанию (25). |
| Протокол интерфейса открытой платформы веб-приложений (WOPI) | Теперь вы можете переименовывать файлы, создавать новые файлы и обмениваться файлами из iframe WOPI на странице браузера. | нет данных |
Подробное описание характеристик
В этом разделе представлены подробные описания новых и обновленных функций SharePoint Server 2016.
Службы Access плюс клиент и сервер Access
Следующие новые функции Access доступны при развертывании служб Access в SharePoint Server 2016:
Центр администрирования больше не предоставляется на всех серверах по умолчанию
Центр администрирования SharePoint Server 2016 теперь по умолчанию предоставляется на первом сервере в ферме при использовании мастера настройки продуктов SharePoint. По умолчанию центр администрирования не инициализируется на дополнительных серверах в ферме.
По умолчанию центр администрирования не инициализируется на дополнительных серверах в ферме.
Вы можете инициализировать или отменить инициализацию центра администрирования на отдельных серверах в ферме, независимо от роли сервера, используя следующие методы:
Страница Services on Server в Central Administration > System Settings
Командлеты Microsoft PowerShell:
Операция
psconfig.exe -cmd adminvsМастер настройки продуктов SharePoint
Примечание
Состояние центра администрирования не влияет на то, считается ли сервер совместимым с MinRole.Правило работоспособности MinRole не будет пытаться инициализировать или отменить инициализацию центра администрирования.
Функции соответствия
Политика удаления документов позволяет удалять документы на сайтах пользователей OneDrive через определенные промежутки времени. Политика хранения на месте позволяет администраторам сохранять документы, электронную почту и другие файлы.
Политика хранения на месте позволяет администраторам сохранять документы, электронную почту и другие файлы.
Дополнительные сведения см. в разделе Обзор политик удаления документов.
Доступ к библиотеке документов
Следующие функции теперь доступны для работы в библиотеках документов SharePoint Server 2016:
Ориентиры на странице упрощают навигацию, а альтернативный текст улучшен для всех основных навигационных ссылок.
Сочетания клавиш предназначены для следующих задач работы с документами:
Улучшения фокуса, такие как сохранение фокуса на предшествующих элементах и захват фокуса.
Объявления о ходе загрузки.
Объявления для имен файлов и типов файлов при просмотре списков папок и файлов.
Улучшено чтение выноски.
Исправлено использование проблем с цветом для переключателя видов.
Обновления справочной документации.

Зашифрованные соединения
Когда вы настраиваете привязку SSL в диспетчере информационных служб Интернета (IIS) для размещения веб-приложения, SharePoint использует шифрование соединения TLS 1.2, если ваше клиентское приложение поддерживает его. SharePoint также поддерживает шифрование соединения TLS 1.2 при подключении к другим системам, например при обходе веб-сайтов.
Примечание
В SSL 3 обнаружена уязвимость безопасности.0, который может позволить злоумышленнику расшифровать данные. В целях повышения безопасности некоторые функции SharePoint теперь по умолчанию отключают шифрование соединения SSL 3.0, а также некоторые алгоритмы шифрования (например, RC4) с известными недостатками. SharePoint отключает шифрование соединения SSL 3.0 по умолчанию для некоторых, но не для всех функций. Чтобы убедиться, что SSL 3.0 отключен для всех функций, вы должны отключить его в Windows, отредактировав реестр Windows. Дополнительные сведения см. в разделах «Отключение SSL 3.0 в Windows для серверного программного обеспечения» и «Для клиентского программного обеспечения», обходные пути в рекомендациях Microsoft по безопасности 3009008.
в разделах «Отключение SSL 3.0 в Windows для серверного программного обеспечения» и «Для клиентского программного обеспечения», обходные пути в рекомендациях Microsoft по безопасности 3009008.
Быстрое создание коллекции веб-сайтов
Эта новая функция предоставляет шаблоны, которые работают на том же уровне, что и SQL Server, что сокращает количество циклов обмена, необходимых между серверами SharePoint и SQL. Используйте командлеты SPSiteMaster Microsoft PowerShell для быстрого создания сайтов и семейств сайтов.
Имена файлов — расширенная поддержка специальных символов
SharePoint исторически блокировал имена файлов, которые включали символы и , ~ , { и } , имена файлов, содержащие GUID , имена файлов с начальными точками и имена файлов длиннее 128 символов. .Эти ограничения сняты в SharePoint Server 2016 и теперь доступны для использования.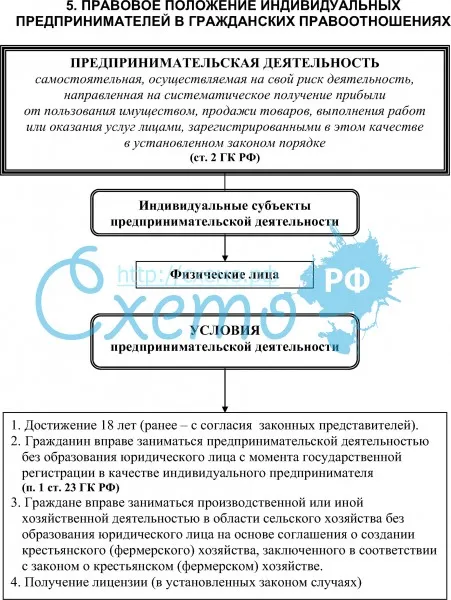
Важно
Запрещенные символы, такие как % и # , по-прежнему не допускаются в именах файлов. Имена файлов страниц, таких как вики-страницы, не могут содержать следующие символы: » # % * : < > ? \ / | и не могут начинаться с ведущей точки (точки).
В SharePoint Server 2016 доступны новые гибридные функции, позволяющие использовать гибридные решения.
Гибридные сайты
Функции гибридных сайтов позволяют вашим пользователям иметь интегрированный опыт при использовании SharePoint Server и SharePoint на сайтах Microsoft 365:
Пользователи могут следить за SharePoint Server и SharePoint на сайтах Microsoft 365 и просматривать их в одном списке.
Пользователи имеют один профиль в Office 365, где хранится вся информация их профиля.
Дополнительные сведения см. в разделе Гибридные сайты и поиск SharePoint.
Гибридный OneDrive
Функции гибридных сайтовиспользуются вместе с Hybrid OneDrive (представлено в SharePoint Server 2013 с пакетом обновления 1 (SP1)):
Облачный гибридный поиск
Облачный гибридный поиск — это новая альтернатива гибридному поисковому решению. С облачным гибридным поиском:
Вы индексируете весь просканированный контент, включая локальный контент, в индекс поиска в Office 365. Вы можете настроить сканер в SharePoint Server 2016 для обхода одних и тех же источников контента и использования тех же соединителей поиска в Office SharePoint Сервер 2007, SharePoint Server 2010 и SharePoint Server 2013.
Когда пользователи запрашивают ваш поисковый индекс в Office 365, они получают унифицированные результаты поиска как из локального содержимого, так и из содержимого Office 365.
Дополнительные сведения о гибридном поиске в облаке см. в общедоступной программе гибридного поиска в облаке Майкрософт при подключении к Microsoft Office.
в общедоступной программе гибридного поиска в облаке Майкрософт при подключении к Microsoft Office.
Дополнительные сведения см. в разделе Планирование гибридного OneDrive.
Дополнительные сведения о гибридных решениях, доступных сегодня, см. в Центре гибридных решений SharePoint.
Благодаря этой новой возможности вы можете:
Поиск конфиденциального контента в SharePoint Server 2016, SharePoint в Microsoft 365 и OneDrive.
Использование 51 встроенный тип конфиденциальной информации (кредитные карты, номера паспортов, номера социального страхования и т. д.).
Чтобы обнаружить конфиденциальный контент, относящийся к общим отраслевым нормам, из центра обнаружения электронных данных SharePoint, в семействе веб-сайтов обнаружения электронных данных выберите DLP Queries , определите нарушающие документы и экспортируйте отчет.
Включите Политики защиты от потери данных из семейства веб-сайтов Центра политики соответствия, чтобы уведомлять конечных пользователей и администраторов, когда документы с конфиденциальной информацией хранятся в SharePoint, и автоматически защищать документы от неправомерного совместного использования.

Сведения о настройке и использовании этой функции задокументированы в SharePoint и Microsoft 365. Дополнительные сведения см. в статье:
.Предварительный просмотр изображений и видео
В SharePoint Server 2016 при публикации изображений и видео в библиотеке документов можно просмотреть предварительный просмотр, наведя указатель мыши на изображение или видео или выбрав их.
Управление правами на доступ к информации
Дополнительные сведения см. в статьях Защита и синхронизация с управлением правами на доступ к данным в OneDrive и Применение управления правами на доступ к списку или библиотеке.
Поддержка больших файлов
Предыдущие версии SharePoint не поддерживали загрузку и загрузку файлов размером более 2047 МБ. SharePoint Server 2016 теперь позволяет загружать или скачивать файлы большего размера. Вы можете настроить желаемый максимальный размер файла для каждого веб-приложения в ферме SharePoint.
Топология фермы MinRole
Роль сервера указывается при создании новой фермы или присоединении сервера к существующей ферме. SharePoint автоматически настраивает службы на каждом сервере в зависимости от роли сервера, оптимизируя производительность фермы на основе этой топологии.Доступно восемь предопределенных ролей сервера, как показано в следующей таблице.
| Роль сервера | Описание |
|---|---|
| Передняя часть | Служебные приложения, службы и компоненты, которые обслуживают запросы пользователей, принадлежат интерфейсным веб-серверам. Эти серверы оптимизированы для низкой задержки. Эти серверы оптимизированы для низкой задержки. |
| Применение | Служебные приложения, службы и компоненты, которые обслуживают внутренние запросы, такие как фоновые задания или запросы обхода контента, принадлежат серверам приложений.Эти серверы оптимизированы для высокой пропускной способности. |
| Распределенный кэш | Служебные приложения, службы и компоненты, необходимые для распределенного кэша, принадлежат серверам распределенного кэша. |
| Поиск | Служебные приложения, службы и компоненты, необходимые для поиска, принадлежат серверам поиска. |
| Пользовательский | Приложения, службы и компоненты пользовательских служб, которые не интегрируются с MinRole, принадлежат настраиваемым серверам.Администратор фермы имеет полный контроль над тем, какие экземпляры службы могут запускаться на серверах, которым назначена роль Custom. MinRole не контролирует, какие экземпляры службы предоставляются для этой роли. MinRole не контролирует, какие экземпляры службы предоставляются для этой роли. |
| Ферма с одним сервером | Служебные приложения, службы и компоненты, необходимые для фермы с одним компьютером, относятся к ферме с одним сервером. Односерверная ферма предназначена для разработки, тестирования и очень ограниченного производственного использования. Ферма SharePoint с ролью фермы с одним сервером не может иметь в ферме более одного сервера SharePoint. Важно: Режим автономной установки больше недоступен в SharePoint Server 2016. Роль фермы с одним сервером заменяет режим автономной установки, доступный в предыдущих выпусках SharePoint Server. В отличие от автономной установки, администратор SharePoint должен отдельно установить и подготовить Microsoft SQL Server для SharePoint. Администратор SharePoint также должен настроить службы и веб-приложения фермы SharePoint либо вручную, либо с помощью мастера настройки фермы.  |
| Внешний интерфейс с распределенным кэшем | Общая роль, объединяющая роли внешнего интерфейса и распределенного кэша на одном сервере. Примечание. Эта общая роль была представлена в ноябрьском общедоступном обновлении для SharePoint Server 2016 (пакет дополнительных компонентов 1). |
| Приложение с поиском | Общая роль, объединяющая роли приложения и поиска на одном сервере. Примечание. Эта общая роль была представлена в ноябрьском общедоступном обновлении для SharePoint Server 2016 (пакет дополнительных компонентов 1). |
Дополнительные сведения о функции MinRole см. в статьях Обзор ролей сервера MinRole в SharePoint Server 2016 и Планирование развертывания сервера MinRole в SharePoint Server 2016.
Мобильный опыт
Когда вы используете мобильное устройство для доступа к домашней странице сайта группы SharePoint Server 2016, вы можете нажимать плитки или ссылки на экране для навигации по сайту. Вы также можете переключиться с представления для мобильных устройств на представление для ПК, в котором страницы сайта отображаются так, как они видны на клиентском компьютере. Этот вид также поддерживает сенсорное управление.
Вы также можете переключиться с представления для мобильных устройств на представление для ПК, в котором страницы сайта отображаются так, как они видны на клиентском компьютере. Этот вид также поддерживает сенсорное управление.
Новые элементы управления для работы с OneDrive
Вы можете выбрать элемент управления, чтобы создавать новые документы Office, загружать файлы, синхронизировать файлы для автономного использования и делиться своими файлами. Дополнительные сведения см. в разделе «Простые элементы управления» в блоге OneDrive.
Открытый формат документа (ODF) доступен для библиотек документов
Формат открытого документа (ODF) позволяет создавать новые файлы в библиотеке документов и сохранять их как файлы ODF, чтобы пользователи могли редактировать новый файл с помощью выбранной ими программы.Дополнительные сведения см. в разделе Установка формата открытого документа (ODF) в качестве шаблона файла по умолчанию для библиотеки.
Проектный сервер 2016
Project Server 2016 для SharePoint Server 2016 имеет множество новых возможностей и функций, в том числе:
Занятость ресурсами : теперь менеджеры проектов могут запрашивать необходимые ресурсы у менеджеров ресурсов для завершения своих проектов.
Кроме того, менеджеры ресурсов могут использовать новую функцию тепловой карты, чтобы увидеть, на что ресурсы тратят свое время.Несколько временных шкал : Менеджеры проектов и портфелей теперь могут создавать более подробные временные шкалы, которые отображают несколько временных шкал в одном представлении.
Более простое администрирование : Project Server теперь имеет возможности многопользовательского хранилища и объединяет хранилище данных с SharePoint. Это значительно снижает накладные расходы на ИТ за счет отказа от выделенной базы данных Project Server и улучшения возможностей резервного копирования и восстановления.
Производительность и масштабирование облачного класса : Многие улучшения производительности и масштабируемости, которые были добавлены в Project Online, также были добавлены в Project Server 2016.
 Кроме того, менеджеры ресурсов могут использовать новую функцию тепловой карты, чтобы увидеть, на что ресурсы тратят свое время.
Кроме того, менеджеры ресурсов могут использовать новую функцию тепловой карты, чтобы увидеть, на что ресурсы тратят свое время. Дополнительные сведения см. в статье Новые возможности Project Server 2016 Preview для ИТ-специалистов.
Важно
Project Server 2016 устанавливается вместе с SharePoint Server 2016 Enterprise, но лицензируется отдельно. Дополнительные сведения о лицензировании Project Server см. в разделе Лицензирование проекта.
SQL Server 2016 CTP 3.1 теперь доступен. Теперь вы можете загрузить SQL Server 2016 CTP 3.1, чтобы использовать надстройку Power Pivot для SharePoint. Вы также можете использовать Power View, установив службы отчетов SQL Server (SSRS) в режиме интеграции с SharePoint и интерфейсную надстройку SSRS с установочного носителя SQL Server.
Загрузите SQL Server 2016 CTP 3.1 из Центра загрузки Майкрософт.
Следующие функции бизнес-аналитики SharePoint Server 2016 доступны при обновлении до SQL Server 2016 CTP 3.1:
Для получения дополнительной информации загрузите новый технический документ «Развертывание SQL Server 2016 PowerPivot и Power View в SharePoint 2016». Для получения подробной информации о настройке и развертывании бизнес-аналитики в ферме SharePoint Server 2016 с несколькими серверами загрузите статью Развертывание SQL Server 2016 PowerPivot и Power View в многоуровневой ферме SharePoint 2016.
Для получения подробной информации о настройке и развертывании бизнес-аналитики в ферме SharePoint Server 2016 с несколькими серверами загрузите статью Развертывание SQL Server 2016 PowerPivot и Power View в многоуровневой ферме SharePoint 2016.
Усовершенствования службы диспетчера запросов
Диспетчер запросов SharePoint теперь предоставляет роли сервера, показанные в следующем списке, для поддержки сценариев регулирования и маршрутизации:
Применение
Распределенный кэш
Передняя часть
Кроме того, служба диспетчера запросов больше не будет препятствовать отображению сайтов, если служба включена, а правила маршрутизации не определены.
Обмен
В следующем списке показаны улучшения общего доступа, доступные для SharePoint Server 2016:
SharePoint Search поддерживает индексирование до 500 миллионов элементов на одно приложение Search Server. Дополнительные сведения см. в статье Обзор архитектуры поиска в SharePoint Server. Сведения об облачном гибридном поиске SharePoint см. в статье Сведения об облачном гибридном поиске для SharePoint.
Дополнительные сведения см. в статье Обзор архитектуры поиска в SharePoint Server. Сведения об облачном гибридном поиске SharePoint см. в статье Сведения об облачном гибридном поиске для SharePoint.
Упрощенная конфигурация SSL для сайта центра администрирования
Мы упростили процесс настройки центра администрирования для использования привязок SSL.Следующие параметры команды теперь доступны для использования:
New-SPCentralAdministration -Port-SecureSocketsLayer Set-SPCentralAdministration -Port-SecureSocketsLayer Psconfig.exe -cmd adminvs -port-ssl
Вы должны назначить сертификат сервера для веб-сайта IIS центра администрирования с помощью инструментов администрирования IIS.Веб-приложение центра администрирования не будет доступно, пока вы не сделаете это.
Если вы укажете порт 443, он автоматически создаст привязку SSL вместо привязки HTTP, даже если вы не включите параметры SecureSocketsLayer или SSL .
Общедоступный URL-адрес AAM центра администрирования будет автоматически обновлен для использования соответствующей схемы протокола, имени сервера и номера порта.
Обновления семейства веб-сайтов
Существует три варианта обновления семейств веб-сайтов.Дополнительные сведения см. в статье Обновление семейства веб-сайтов до SharePoint Server 2016.
Шифрование SMTP-соединения
В следующем списке показаны требования SharePoint 2016, необходимые для согласования шифрования соединения с SMTP-сервером:
STARTTLS должен быть включен на SMTP-сервере.
SMTP-сервер должен поддерживать протокол TLS 1.0, TLS 1.1 или TLS 1.2.
Важно
Протоколы SSL 2.0 и SSL 3.0 не поддерживаются.
На SMTP-сервере должен быть установлен сертификат сервера.
Сертификат сервера должен быть действительным. Как правило, это означает, что имя сертификата сервера должно совпадать с именем SMTP-сервера, предоставленным SharePoint.
Сертификат сервера также должен быть выдан центром сертификации, которому доверяет сервер SharePoint.В SharePoint необходимо настроить шифрование SMTP-соединения.
 Сертификат сервера также должен быть выдан центром сертификации, которому доверяет сервер SharePoint.
Сертификат сервера также должен быть выдан центром сертификации, которому доверяет сервер SharePoint. Чтобы настроить SharePoint на постоянное использование шифрования SMTP-подключения, откройте веб-сайт центра администрирования SharePoint и перейдите к Параметры системы > Настройте параметры исходящей электронной почты и установите в раскрывающемся меню Использовать шифрование подключения TLS значение Да. .Чтобы настроить SharePoint на постоянное использование шифрования SMTP-подключения в Microsoft PowerShell, используйте командлет Set-SPWebApplication без параметра DisableSMTPEncryption . Например:
$WebApp = Get-SPWebApplication -IncludeCentralAdministration | ? { $_.IsAdministrationWebApplication -eq $true}
Set-SPWebApplication -Identity $WebApp -SMTPServer smtp. internal.contoso.com -OutgoingEmailAddress [email protected] -ReplyToEmailAddress sharepoint@contoso.ком
 internal.contoso.com -OutgoingEmailAddress
internal.contoso.com -OutgoingEmailAddress Чтобы настроить SharePoint так, чтобы он никогда не использовал шифрование SMTP-подключения в центре администрирования SharePoint, перейдите к Системные настройки > Настройте параметры исходящей электронной почты и установите в раскрывающемся меню Использовать шифрование подключения TLS значение Нет . Чтобы настроить SharePoint так, чтобы он никогда не использовал шифрование SMTP-подключения в PowerShell, используйте командлет Set-SPWebApplication с параметром DisableSMTPEncryption . Например:
$WebApp = Get-SPWebApplication -IncludeCentralAdministration | ? { $_.IsAdministrationWebApplication -eq $true }
Set-SPWebApplication -Identity $WebApp -SMTPServer smtp.internal.contoso.com -DisableSMTPEncryption -OutgoingEmailAddress sharepoint@contoso. com -ReplyToEmailAddress
[email protected]
 com -ReplyToEmailAddress
com -ReplyToEmailAddress
Примечание
Если SharePoint настроен на использование шифрования SMTP-соединения, он будет отправлять сообщения электронной почты только в том случае, если успешно согласовал шифрование соединения с SMTP-сервером. Он не будет отступать и отправлять сообщения электронной почты в незашифрованном виде в случае сбоя согласования шифрования соединения.Если SharePoint не настроен на использование шифрования SMTP-подключения, он всегда будет отправлять сообщения электронной почты в незашифрованном виде, даже если SMTP-сервер поддерживает шифрование подключения. > Использование шифрования соединения SMTP не включает аутентификацию SMTP. SMTP-запросы всегда отправляются анонимно.
Просмотр папок сайта
Дополнительные сведения см. в разделе «Папки сайта» в блоге OneDrive.
Закрепление страницы сайта
Теперь вы можете закреплять сайты, которые видите на странице сайтов. Закрепленный сайт отображается вверху списка сайтов, на которые вы подписаны.
Набор навигации по темам
Теперь вы можете применять темы к своей навигации Suite.
Использовать порты SMTP, отличные от портов по умолчанию (25)
Чтобы настроить SharePoint для использования порта SMTP, отличного от порта по умолчанию, откройте центр администрирования SharePoint, перейдите к Системные настройки > Настройка параметров исходящей электронной почты и установите порт SMTP-сервера на номер порта вашего SMTP-сервера. Чтобы настроить SharePoint для использования порта SMTP, отличного от порта по умолчанию в PowerShell, используйте командлет Set-SPWebApplication с параметром SMTPServerPort
$WebApp = Get-SPWebApplication -IncludeCentralAdministration | ? { $_.IsAdministrationWebApplication -eq $true}
Set-SPWebApplication -Identity $WebApp -SMTPServer smtp.internal.contoso.com -SMTPServerPort 587 -OutgoingEmailAddress sharepoint@contoso. com -ReplyToEmailAddress
sh [email protected]
 com -ReplyToEmailAddress
sh
com -ReplyToEmailAddress
sh Что такое SharePoint?
Penicillium Sp. | Виноградарство и виноделие
Род/вид: Penicillium Sp.
Классификация: Аскомицет; анаморф ( Penicillium часто упоминается как Deuteromycetes)
Морфология:
- Клетка: многоклеточная, эллипсовидная
- Спора: Конидия; фиалиди
- Зигота: размножается бесполым путем, зиготы отсутствуют
- Аск: нет данных
- Жидкий рост: пленка, биопленка
Penicillum spp. являются нитчатыми грибами. Имеют разветвленные конидиоспоры. Конидии округлые, одноклеточные.Penicillium размножается бесполым путем. Виды классифицируют по способу образования конидий. У некоторых видов конидии рождаются на фиалидиях. У других конидиофор несет метуллы, из которых рождаются фиалидии. У других видов конидиофор может разветвляться до образования метул.
Физиологические признаки:
- Сапрофит, питательные вещества из мертвых и разложившихся веществ
- Плесень Penicillium ингибирует образование бактериальной биопленки частично за счет продукции пенициловой кислоты и патулина
- Сухие споры легко распространяются по воздуху, а также переносятся водой
- Производитель токсинов и аллергенных соединений
Экологические характеристики:
- Вездесущий
- Почва, злаки, силос, хлеб, лук, чеснок, сыр, орехи, фрукты и другие продукты
- Лишайники (состоящие из двух разных видов: гриба и либо цианобактерии, либо зеленой водоросли)
- Penicillium spp. .штаммы более распространены в регионах с умеренным климатом
Отличительные признаки:
- Penicillium spp. . изначально белые, со временем становятся сине-зелеными, серо-зелеными, оливково-серыми, желтыми или розоватыми
- Многоклеточные грибы состоят из нитей, называемых гифами
- Гифы могут содержать внутренние поперечные стенки, называемые перегородками , которые делят гифы на отдельные клетки
- хитин/глюкановая стенка
- Споры (конидии) образуются сухими цепочками из кончиков фиалидов, с самой молодой спорой в основании цепочки, и почти всегда зеленые
Роль в вине:
- Массивное негативное воздействие на органолептику вина
- Потенциальное увеличение титруемой кислотности
- Споры Penicillium содержат микотоксины (патулин, цитринин, рокефортин, охратоксин А – «OTA»), вызывающие различные симптомы аллергии и заболевания
- OTA: Международное агентство по изучению рака классифицировало OTA в группе 2B как возможный канцероген для человека
- Грибы могут изменять химический состав ягод и влиять на рост дрожжей во время спиртового брожения, что приводит к изменению цвета и вкуса вина
Чувствительность:
- SO2: Х
- Сорбат: X
- DMDC: Х
- рН: Х
- Кислоты:X
- Этанол: Х
- Анаэробиоз: №
- Тепло: > 55°C
Каталожные номера:
Торелли, Э. et al, 2006. Штаммы Penicillium spp., продуцирующие охратоксин А. выделен из винограда, используемого для производства вин «пассито». International Journal of Food Microbiology 106 (2006) 307 – 312
et al, 2006. Штаммы Penicillium spp., продуцирующие охратоксин А. выделен из винограда, используемого для производства вин «пассито». International Journal of Food Microbiology 106 (2006) 307 – 312
Особенности генома умеренно галофильных продуцентов полигидроксиалканоатов Yangia sp. ЦКБ-ММ3 | Экологический микробиом
Dai X, Wang B-J, Yang Q-X, Jiao N-Z, Liu S-J. Yangia pacifica род. ноябрь, сп. nov., новый член клады Roseobacter из прибрежных отложений Восточно-Китайского моря.Int J Syst Evol Microbiol. 2006; 56: 529–33.
КАС Статья пабмед Google Scholar
Пухалте М.Дж., Лусена Т., Рувира М.А., Арахал Д.Р., Масиан М.С. Семейство Rhodobacteraceae . В: Розенберг Э., Делонг Э. Ф., Стакебрандт Э., Лори С., Томпсон Ф., редакторы. Прокариоты-альфапротеобактерии и бетапротеобактерии, том. 4. Берлин: Спрингер; 2014. с. 439–512.
Глава Google Scholar

Бьюкен А., Гонсалес Х.М., Моран М.А. Обзор морской линии Roseobacter . Appl Environ Microbiol. 2005; 71: 5665–77.
КАС Статья пабмед ПабМед Центральный Google Scholar
Van-Thuoc D, Huu-Phong T, Minh-Khuong D, Hatti-Kaul R. Получение поли(3-гидроксибутират- со -3-гидроксивалерата) умеренным галофилом Yangia sp. ND199 с использованием глицерина в качестве источника углерода.Заявл. Биохим Биотехнолог. 2015;175:3120–32.
КАС Статья пабмед Google Scholar
Хань Дж., Хоу Дж., Чжан Ф., Ай Г., Ли М., Цай С., Лю Х., Ван Л., Ван З., Чжан С. и др. Множественные пути снабжения пропионил-коферментом А для производства биопластического поли(3-гидроксибутират- со -3-гидроксивалерата) в Haloferax mediterranei . Appl Environ Microbiol. 2013;79:2922–31.
КАС Статья пабмед ПабМед Центральный Google Scholar

Валентин Х.Ф., Деннис Д. Метаболический путь образования поли(3-гидроксибутирата- со -3-гидроксивалерата) в Nocardia corallina : инактивация mutB путем хромосомной интеграции гена устойчивости к канамицину. Appl Environ Microbiol. 1996; 62: 372–9.
КАС пабмед ПабМед Центральный Google Scholar
Son H, Lee S. Биосинтез поли(3-гидроксибутирата- со -3-гидроксивалерата) из структурно несвязанных одиночных источников углерода вновь выделенным штаммом Pseudomonas sp.ЭЛ-2. Биотехнологическая лат. 1996; 18:1217–22.
КАС Статья Google Scholar
Haywood GW, Anderson AJ, Williams DR, Dawes EA, Ewing DF. Накопление поли(гидроксиалканоатного) сополимера, содержащего в основном 3-гидроксивалерат, из простых углеводных субстратов Rhodococcus sp. NCIMB 40126. Int J Biol Macromol. 1991; 13:83–88.
КАС Статья пабмед Google Scholar

Chen Q, Wang Q, Wei G, Liang Q, Qi Q. Производство в Escherichia coli поли(3-гидроксибутирата- со -3-гидроксивалерата) с различным составом мономеров из несвязанных источников углерода. Appl Environ Microbiol. 2011;77:4886–93.
КАС Статья пабмед ПабМед Центральный Google Scholar
Tsuge T. Метаболические улучшения и использование недорогих источников углерода в микробном производстве полигидроксиалканоатов.J Biosci Bioeng. 2002; 94: 579–84.
КАС Статья пабмед Google Scholar
Джусофф К. Мангровые леса Малайзии и их значение для прибрежной морской среды. Pol J Environ Stud. 2013;22:979–1005.
Google Scholar
Инь Дж, Чен Дж-К, Ву Кью, Чен Г-К. Галофилы, грядущие звезды промышленной биотехнологии. Биотехнология Adv. 2015; 33:1433–42.
КАС Статья пабмед Google Scholar

Динеш Б., Лау Н.С., Фурусава Г., Ким С.В., Тейлор Т.Д., Фун С.И., Шу-Чьен А.С. Сравнительный анализ генома новых штаммов, подобных Mangrovimonas , выделенных из отложений эстуарных мангровых зарослей, выявил гены утилизации ксилана и арабинана. Мар Геномикс. 2016;25:115–21.
Артикул пабмед Google Scholar
Фурусава Г., Лау Н.С., Шу-Чьен А.С., Джая-Рам А., Амирул А.А.А. Идентификация путей биосинтеза полиненасыщенных жирных кислот и дитерпеноидов из проекта генома Aureispira sp. ЦКБ-QB1. Мар Геномикс. 2015;19:39–44.
Артикул пабмед Google Scholar
Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW. EzTaxon: веб-инструмент для идентификации прокариот на основе последовательностей генов 16S рибосомной РНК.Int J Syst Evol Microbiol. 2007; 57: 2259–61.
КАС Статья пабмед Google Scholar

Lau N-S, Tsuge T, Sudesh K. Образование нового полигидроксиалканоата, содержащего мономер 3-гидрокси-4-метилвалерат, в Burkholderia sp. Приложение Microbiol Biotechnol. 2011; 89: 1599–609.
КАС Статья пабмед Google Scholar
Чин К.С., Александр Д.Х., Маркс П., Кламмер А.А., Дрейк Дж., Хайнер С., Клам А., Коупленд А., Хаддлстон Дж., Эйхлер Э.Е. и др.Негибридные, готовые сборки микробного генома из давно прочитанных данных секвенирования SMRT. Нат Мет. 2013;10:563–9.
КАС Статья Google Scholar
Азиз Р.К., Бартелс Д., Бест А.А., ДеДжонг М., Диз Т., Эдвардс Р.А., Формсма К., Гердес С., Гласс Э.М., Кубал М. и др. Сервер RAST: быстрые аннотации с использованием технологии подсистем. Геномика BMC. 2008; 9:75.
Артикул пабмед ПабМед Центральный Google Scholar

Lowe ТМ, Эдди С.Р. tRNAscan-SE: программа для улучшенного обнаружения генов транспортной РНК в геномной последовательности. Нуклеиновые Кислоты Res. 1997; 25:0955–64.
КАС Статья Google Scholar
Dyrløv Bendtsen J, Nielsen H, von Heijne G, Brunak S. Улучшенное предсказание сигнальных пептидов: SignalP 3.0. Дж Мол Биол. 2004; 340:783–95.
Артикул Google Scholar
Крог А., Ларссон Б., фон Хейне Г., Зоннхаммер Э.Л. Прогнозирование топологии трансмембранных белков с помощью скрытой марковской модели: приложение к полным геномам. Дж Мол Биол. 2001; 305: 567–80.
КАС Статья пабмед Google Scholar
Grissa I, Vergnaud G, Pourcel C. CRISPRFinder: веб-инструмент для выявления сгруппированных коротких палиндромных повторов с регулярными интервалами. Нуклеиновые Кислоты Res. 2007; 35: W52–57.
2007; 35: W52–57.
Артикул пабмед ПабМед Центральный Google Scholar
Gonzalez JM, Covert JS, Whitman WB, Henriksen JR, Mayer F, Scharf B, Schmitt R, Buchan A, Fuhrman JA, Kiene RP, et al. Silicibacter pomeroyi sp. ноябрь и Roseovarius nubinhibens sp. nov., диметилсульфониопропионат-деметилирующие бактерии из морской среды. Int J Syst Evol Microbiol. 2003;53:1261–9.
КАС Статья пабмед Google Scholar
Пукалл Р., Бунтефус Д., Фрулинг А., Роде М., Кроппенштедт Р.М., Бургхардт Дж., Лебарон П., Бернард Л., Штакебрандт Э. Sulfitobacter mediterraneus sp. nov., новый сульфитокисляющий представитель α-протеобактерий. Int J Syst Bacteriol. 1999; 49: 513–9.
КАС Статья пабмед Google Scholar
Сорокин Д.Ю. Sulfitobacter pontiacus род. ноябрь, сп. ноябрь — новая гетеротрофная бактерия из Черного моря, специализирующаяся на окислении сульфитов. Микробиология. 1995; 64: 354–65.
ноябрь, сп. ноябрь — новая гетеротрофная бактерия из Черного моря, специализирующаяся на окислении сульфитов. Микробиология. 1995; 64: 354–65.
КАС Google Scholar
Сове В., Бруно С., Беркс Б.К., Хеммингс А.М. Комплекс SoxYZ несет промежуточные продукты серного цикла на пептидном качающемся рычаге. Дж. Биол. Хим. 2007; 282:23194–204.
КАС Статья пабмед Google Scholar
Kilmartin JR, Maher MJ, Krusong K, Noble CJ, Hanson GR, Bernhardt PV, Riley MJ, Kappler U. Взгляд на структуру и функцию активного центра цитохромов SoxAX. Дж. Биол. Хим. 2011; 286:24872–81.
КАС Статья пабмед ПабМед Центральный Google Scholar
Сове В., Роверси П., Лит К.Дж., Гарман Э.Ф., Антробус Р., Леа С.М., Беркс Б.К. Механизм гидролиза связи сера-сера на основе кристаллической структуры тиосульфогидролазы SoxB. Дж. Биол. Хим. 2009; 284:21707–18.
Дж. Биол. Хим. 2009; 284:21707–18.
КАС Статья пабмед ПабМед Центральный Google Scholar
Цандер У., Фауст А., Клинк Б.Ю., де Санктис Д., Панджикар С., Квентмайер А., Бардишевский Ф., Фридрих К.Г., Шайдиг А.Я. Структурные основы окисления связанной с белком серы молибдогемоферментной сульфандегидрогеназой SoxCD серного цикла.Дж. Биол. Хим. 2011; 286:8349–60.
КАС Статья пабмед Google Scholar
Уильямс Д.Р., Андерсон А.Дж., Доус Э.А., Юинг Д.Ф. Производство сополиэфира 3-гидроксимасляной кислоты и 3-гидроксивалериановой кислоты из янтарной кислоты с помощью Rhodococcus ruber : соображения биосинтеза. Приложение Microbiol Biotechnol. 1994;40:717–23.
КАС Статья Google Scholar
Stubbe J, Tian J. Полигидроксиалканоатный (PHA) гемостаз: роль PHA-синтазы. Nat Prod Rep. 2003; 20: 445–57.
Nat Prod Rep. 2003; 20: 445–57.
КАС Статья пабмед Google Scholar
Rehm КНБК. Полиэфирсинтазы: природные катализаторы для пластмасс. Биохим Дж. 2003; 376:15–33.
КАС Статья пабмед ПабМед Центральный Google Scholar
Джендроссек Д., Хэндрик Р.Микробная деградация полигидроксиалканоатов. Анну Рев Микробиол. 2002; 56: 403–32.
КАС Статья пабмед Google Scholar
Neumann L, Spinozzi F, Sinibaldi R, Rusticelli F, Pötter M, Steinbüchel A. Связывание основного фазина PhaP1 из Ralstonia eutropha h26 с гранулами поли(3-гидроксибутирата). J Бактериол. 2008;190:2911–9.
КАС Статья пабмед ПабМед Центральный Google Scholar
Maehara A, Taguchi S, Nishiyama T, Yamane T, Doi Y. Белок-репрессор PhaR регулирует синтез полигидроксиалканоатов (PHA) посредством прямого взаимодействия с PHA. J Бактериол. 2002; 184:3992–4002.
Белок-репрессор PhaR регулирует синтез полигидроксиалканоатов (PHA) посредством прямого взаимодействия с PHA. J Бактериол. 2002; 184:3992–4002.
КАС Статья пабмед ПабМед Центральный Google Scholar
Woese CR, Kandler O, Wheelis ML. На пути к естественной системе организмов: предложение для доменов Archaea, Bacteria и Eucarya. Proc Natl Acad Sci U S A.1990; 87: 4576–9.
КАС Статья пабмед ПабМед Центральный Google Scholar
Гаррити ГМ, Белл Дж. А., Лилберн Т. Г. Тип XIV. Протеобактерии фил. ноябрь В: Garrity GM, Brenner DJ, Krieg NR, Staley JT, редакторы. Руководство Берджи по систематической бактериологии, том. 2. Второе изд. Нью-Йорк: Спрингер; 2005.
Google Scholar
Гаррити ГМ, Белл Дж. А., Лилберн Т. Г.Класс I. Альфапротеобактерии класс. ноябрь В: Garrity GM, Brenner DJ, Krieg NR, Staley JT, редакторы. Руководство Берджи по систематической бактериологии, том. 2. Второе изд. Нью-Йорк: Спрингер; 2005.
ноябрь В: Garrity GM, Brenner DJ, Krieg NR, Staley JT, редакторы. Руководство Берджи по систематической бактериологии, том. 2. Второе изд. Нью-Йорк: Спрингер; 2005.
Google Scholar
Гаррити ГМ, Белл Дж. А., Лилберн Т. Г. Орден III. Rhodobacterales орд. ноябрь В: Garrity GM, Brenner DJ, Krieg NR, Staley JT, редакторы. Руководство Берджи по систематической бактериологии, том. 2. Второе изд.Нью-Йорк: Спрингер; 2005.
Google Scholar
Гаррити ГМ, Белл Дж. А., Лилберн Т. Г. Семейство I. Rhodobacteraceae сем. ноябрь В: Garrity GM, Brenner DJ, Krieg NR, Staley JT, редакторы. Руководство Берджи по систематической бактериологии, том. 2. Второе изд. Нью-Йорк: Спрингер; 2005.
Google Scholar
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Генная онтология: инструмент для объединения биологии. Консорциум генных онтологий. Нат Жене. 2000; 25:25–9.
Генная онтология: инструмент для объединения биологии. Консорциум генных онтологий. Нат Жене. 2000; 25:25–9.
КАС Статья пабмед ПабМед Центральный Google Scholar
Saitou N, Nei M. Метод объединения соседей: новый метод реконструкции филогенетических деревьев. Мол Биол Эвол. 1987; 4: 406–25.
КАС пабмед Google Scholar
Тамура К., Стечер Г., Петерсон Д., Филипски А., Кумар С.MEGA6: Молекулярно-эволюционный генетический анализ версии 6.0. Мол Биол Эвол. 2013;30:2725–9.
КАС Статья пабмед ПабМед Центральный Google Scholar
Grant JR, Arantes AS, Stothard P. Сравнение тысяч кольцевых геномов с использованием инструмента сравнения CGView. Геномика BMC. 2012;13:202.
КАС Статья пабмед ПабМед Центральный Google Scholar
Arrow McLaren SP представит схему окраски победителя Vuse Design Challenge на улицах Нэшвилла
Arrow McLaren SP представит схему окраски победителя Vuse Design Challenge на улицах Нэшвилла
Дизайн победителя конкурса Vuse Design Challenge Джека ЛаПилуса будет представлен как на первом, так и на втором5 Arrow McLaren SP Chevrolet и Vuse Arrow McLaren SP Chevrolet № 7 на первом Гран-при Big Machine Music City
Arrow McLaren SP и Vuse представляют дизайн, победивший в конкурсе Vuse Design Challenge, который будет представлен как на № 5 Arrow McLaren SP Chevrolet, так и на № 7 Vuse Arrow McLaren SP Chevrolet на предстоящем первом Гран-при Big Machine Music City. в Нэшвилле, Теннесси.
в Нэшвилле, Теннесси.
Давний поклонник Arrow McLaren SP, Джек ЛаПилуса из Астории, Нью-Йорк, представил победившую заявку на конкурс Vuse Design Challenge, черпая вдохновение в ослепительном камуфляже времен Первой мировой войны для своей уникальной окраски.
«Победа в Vuse Design Challenge заставляет меня чувствовать себя на вершине мира. Знать, что мой дизайн будет участвовать в гонках по Нэшвиллу, представляя такую престижную гоночную команду, — это особое и сюрреалистическое чувство», — сказал ЛаПилуса. «Я изучил десятки различных камуфляжей прототипов автомобилей, которые производители используют, чтобы скрыть свои будущие модели, и создал шаблоны, вдохновленные некоторыми из наиболее знаковых дизайнов. Затем я дал ему цветовую комбинацию, которую фанаты McLaren и жители Нью-Йорка, такие как я, узнают и полюбят.Машина может быть «замаскирована», но я уверен, что вы ее заметите».
В дизайне представлены фирменные цвета каждого автомобиля, знакомые поклонникам: № 5 Arrow McLaren SP Chevrolet с цветами папайи и черного цвета, а № 7 Vuse Arrow McLaren SP Chevrolet — цвета папайи и кобальтово-синего цвета.
«Дизайн Джека великолепен и действительно говорит о том, чего мы пытаемся достичь как команда дизайнеров наших гоночных автомобилей в McLaren Racing», — сказала Луиза Макьюэн, директор по бренду и креативу McLaren Racing.«Дизайн должен быть революционным и выделяться, и это именно то, что нужно для автомобилей № 5 и № 7. Было приятно принять участие в конкурсе Vuse Design Challenge вместе с другими судьями и стать свидетелем этой прекрасной возможности для болельщика привнести свой талант и креативность в команду Arrow McLaren SP».
Увидев дизайн Джека, Arrow Electronics восприняла дух конкурса Vuse Design Challenge и представила индивидуальную окраску для всей команды Arrow McLaren SP.
Дизайн Джека был выбран из сотен участников конкурса Vuse Design Challenge, и судьи сочли, что он идеально подходит для команды.Как победитель, он также получит 10 000 долларов призовых и VIP-опыт на гонке в Нэшвилле, что позволит ему впервые увидеть свои работы на улицах на первом Гран-при Big Machine Music City в воскресенье. 8 августа в 17:30 ET на NBCSN.
8 августа в 17:30 ET на NBCSN.
«Я говорю за всю команду, когда говорю, что мы взволнованы, чтобы добраться до Нэшвилла. Вы можете видеть, что наши ключевые партнеры разделяют волнение, так как Vuse Design Challenge собирается создать две новые ливреи для Arrow и Vuse на No.Автомобили № 5 и № 7 соответственно», — сказал президент Arrow McLaren SP Тейлор Кил. «Я знаю, что Джек является большим поклонником Arrow McLaren SP, и он проявил свою страсть с большим творческим потенциалом. Для него будет особенным моментом лично увидеть дизайн в Нэшвилле».
Arrow McLaren SP и Vuse продемонстрировали свою общую страсть к искусству и изобретательность на протяжении всего сезона NTT INDYCAR SERIES, демонстрируя несколько совместных работ с местными художниками, в том числе во время гоночных уик-эндов в Сент-Луисе.Санкт-Петербург, Форт-Уэрт и Индианаполис. Каждый художник создал свой уникальный дизайн, воплощенный в жизнь во время живого рисования выставленного гоночного автомобиля.
Vuse Design Challenge является частью партнерства бренда с Arrow McLaren SP и более широкой расширенной партнерской сделки между BAT, R.J. Конечная материнская группа Reynolds Vapor Company и McLaren Racing в качестве продолжения глобального партнерства с командой McLaren Formula 1, основным партнером которой является BAT.
«Было удивительно видеть, как Vuse Design Challenge воплощается в жизнь и вдохновляет наших потребителей демонстрировать свои творческие способности», — сказал Хорхе Арайя, исполнительный вице-президент и коммерческий директор Reynolds.«Страсть бренда к искусству и инновациям привела к этой уникальной возможности. Мы рады видеть, что дизайн Джека попал на трассу в Нэшвилле».
Поделись новостью!
границ | Геномные особенности устойчивости к высыханию и биосинтеза сахаров у Extremophile Gloeocapsopsis sp. УТЕКС B3054
Введение
Пустыня Атакама — самая сухая теплая пустыня на Земле (Houston and Hartley, 2003; Hartley et al. , 2005). Расположенная в северной части Чили, она в течение многих лет считалась стерильной территорией, неспособной дать убежище ни одному живому организму (McKay et al., 2003; Наварро-Гонсалес и др., 2003). Недавно мы узнали, что в пустыне Атакама случайные поступления воды из прибрежного тумана и росы поддерживают скудную микробную жизнь, которая процветает в характерных для нее экстремальных условиях окружающей среды (Houston and Hartley, 2003; McKay et al., 2003).
, 2005). Расположенная в северной части Чили, она в течение многих лет считалась стерильной территорией, неспособной дать убежище ни одному живому организму (McKay et al., 2003; Наварро-Гонсалес и др., 2003). Недавно мы узнали, что в пустыне Атакама случайные поступления воды из прибрежного тумана и росы поддерживают скудную микробную жизнь, которая процветает в характерных для нее экстремальных условиях окружающей среды (Houston and Hartley, 2003; McKay et al., 2003).
Микробная жизнь разработала множество физических и молекулярных стратегий для преодоления высокой солнечной радиации и высоких температур, а также для максимизации эффективности использования небольшого количества доступной воды.Действительно, большая часть микробной жизни в пустынях так или иначе связана со скалами, развиваясь либо внутри них, либо под ними (Chan et al., 2012; Pointing and Belnap, 2012; Cowan et al., 2014; Davila et al., 2015; Wierzchos et al. ., 2015). В этих микробных сообществах преобладают первичные продуценты, представленные преимущественно морфологически и метаболически разнообразными цианобактериями (Pointing et al. , 2009; Wong et al., 2010; Wierzchos et al., 2015; Wei et al., 2016). Большинство этих пустынных цианобактерий образуют обильные внеклеточные структуры, что, как считается, составляет как архитектурную, так и метаболическую основу микробного сообщества и его устойчивости к экстремальным условиям окружающей среды (Knowles and Castenholz, 2008; Colica et al., 2014; Росси и Де Филиппис, 2015).
, 2009; Wong et al., 2010; Wierzchos et al., 2015; Wei et al., 2016). Большинство этих пустынных цианобактерий образуют обильные внеклеточные структуры, что, как считается, составляет как архитектурную, так и метаболическую основу микробного сообщества и его устойчивости к экстремальным условиям окружающей среды (Knowles and Castenholz, 2008; Colica et al., 2014; Росси и Де Филиппис, 2015).
В частности, цианобактерии, организованные в пакетообразные структуры, такие как Chroococcidiopsis и Gloeocapsa , доминируют в каменных сообществах, встречающихся в теплых пустынях (Warren-Rhodes et al., 2006; Bahl et al., 2011; Chan et al., 2012). ; Wierzchos et al., 2015; Crits-Christoph et al., 2016). Хотя эти микроорганизмы были идентифицированы с помощью микроскопии и исследований 16S-рДНК, существенным препятствием для прогресса в изучении цианобактерий в целом были трудности, возникающие при получении аксеновых культур, а также наличие большого количества экзополисахарида (ЭПС), препятствующего секвенирование их геномов (Tillett and Neilan, 2000; Chrismas et al. , 2016). Чтобы проиллюстрировать последнее, только 1110 из 76 299 геномов, доступных в базе данных Integrated Microbial Genomes and Microbiomes (IMG/JGI), соответствуют цианобактериям. Только морской род Prochlorococcus концентрирует 55,85% уже имеющихся геномов цианобактерий. Более того, всего три генома соответствуют цианобактериям, выделенным из пустынной среды, и все они нитевидные.
, 2016). Чтобы проиллюстрировать последнее, только 1110 из 76 299 геномов, доступных в базе данных Integrated Microbial Genomes and Microbiomes (IMG/JGI), соответствуют цианобактериям. Только морской род Prochlorococcus концентрирует 55,85% уже имеющихся геномов цианобактерий. Более того, всего три генома соответствуют цианобактериям, выделенным из пустынной среды, и все они нитевидные.
Растущий объем знаний подтверждает фундаментальную роль совместимых растворенных веществ (Hershkovitz et al., 1991; Хилл и др., 1994, 1997; Сакамото и др., 2009 г.; Ёсида и Сакамото, 2009 г.; Klähn and Hagemann, 2011) и EPS (Grilli Caiola et al., 1993, 1996; Hill et al., 1997; Tamaru et al., 2005; Knowles and Castenholz, 2008; Mager and Thomas, 2011) могут влиять на устойчивость к высыханию. у цианобактерий. Однако всесторонний геномный анализ механизмов устойчивости к экстремальному высыханию у одноклеточных цианобактерий все еще отсутствует. В связи с этим мы решили секвенировать и изучить геном Gloeocapsopsis sp. UTEX B3054, одноклеточная цианобактерия, которую мы продемонстрировали, принадлежит к семейству Chroococcaceae и имеет несколько известных культивируемых и секвенированных представителей. Этот штамм был получен путем сортировки клеток из Gloeocapsopsis sp. Культура AAB1, обогащение, первоначально собранное из кварцевой породы в пустыне Атакама и описанное как чрезвычайно устойчивое к высыханию (Azúa-Bustos et al., 2014). Помимо улучшения охвата генома этого семейства цианобактерий, наше исследование было направлено на прогнозирование и анализ геномных механизмов, которые, вероятно, связаны с устойчивостью к высыханию Gloeocapsopsis sp.УТЕКС B3054. В частности, мы сосредоточились на выявлении генетического потенциала и геномных механизмов, которые, вероятно, участвуют в биосинтезе совместимых растворенных веществ и ЭПС, молекул, которые играют ключевую роль в устойчивости микробов к засухе.
UTEX B3054, одноклеточная цианобактерия, которую мы продемонстрировали, принадлежит к семейству Chroococcaceae и имеет несколько известных культивируемых и секвенированных представителей. Этот штамм был получен путем сортировки клеток из Gloeocapsopsis sp. Культура AAB1, обогащение, первоначально собранное из кварцевой породы в пустыне Атакама и описанное как чрезвычайно устойчивое к высыханию (Azúa-Bustos et al., 2014). Помимо улучшения охвата генома этого семейства цианобактерий, наше исследование было направлено на прогнозирование и анализ геномных механизмов, которые, вероятно, связаны с устойчивостью к высыханию Gloeocapsopsis sp.УТЕКС B3054. В частности, мы сосредоточились на выявлении генетического потенциала и геномных механизмов, которые, вероятно, участвуют в биосинтезе совместимых растворенных веществ и ЭПС, молекул, которые играют ключевую роль в устойчивости микробов к засухе.
Материалы и методы
Выделение штамма и выделение ДНК
Штамм, использованный в этом исследовании , Gloeocapsopsis sp. UTEX B3054, был получен из неаксенового Gloeocapsopsis sp. Обогащающая культура AAB1, первоначально выделенная из пустыни Атакама (Azúa-Bustos et al., 2014). Для массового устранения загрязняющих гетеротрофных бактерий одну цианобактериальную клетку сортировали в среду BG11 с использованием сортировщика клеток Influx Mariner Cell Sorter (Cytopeia, Сиэтл, Вашингтон, США). Хлорофиллсодержащие клетки выявляли по красной флуоресценции (692–40 нм; флуоресцентные фильтры указаны здесь по длине волны максимального пропускания и спектральной ширине полосы пропускания), возбуждаемой лазером с длиной волны 488 нм, а запуск основывался на боковом рассеянии света (SSC). ), чтобы исключить нефлуоресцентные клетки.Клон Gloeocapsopsis sp. Культура UTEX B3054 была депонирована в Коллекции культур водорослей UTEX под регистрационным кодом UTEX B3054 и находится в открытом доступе.
UTEX B3054, был получен из неаксенового Gloeocapsopsis sp. Обогащающая культура AAB1, первоначально выделенная из пустыни Атакама (Azúa-Bustos et al., 2014). Для массового устранения загрязняющих гетеротрофных бактерий одну цианобактериальную клетку сортировали в среду BG11 с использованием сортировщика клеток Influx Mariner Cell Sorter (Cytopeia, Сиэтл, Вашингтон, США). Хлорофиллсодержащие клетки выявляли по красной флуоресценции (692–40 нм; флуоресцентные фильтры указаны здесь по длине волны максимального пропускания и спектральной ширине полосы пропускания), возбуждаемой лазером с длиной волны 488 нм, а запуск основывался на боковом рассеянии света (SSC). ), чтобы исключить нефлуоресцентные клетки.Клон Gloeocapsopsis sp. Культура UTEX B3054 была депонирована в Коллекции культур водорослей UTEX под регистрационным кодом UTEX B3054 и находится в открытом доступе.
Далее было проведено несколько обработок с целью механической, химической и ферментативной дестабилизации агрегатов цианобактерий, селективного удаления молекул сахара и устранения устойчивых сопутствующих гетеротрофных бактерий в культуре. Первая обработка была применена к здоровым культурам в середине активного роста и была основана на ранее опубликованном протоколе для клеток, богатых сахаром (Sharma et al., 2002), с несколькими модификациями, подробно описанными на дополнительном рисунке S1. Для обеспечения деградации оставшейся ДНК нецианобактерий осадок клеток цианобактерий, полученный после первой обработки, ресуспендировали в 100 мкл стерильной воды, содержащей четыре единицы DN азы I (Invitrogen), и инкубировали при 37°C в течение 1 час Ферментативную реакцию останавливали при 65°С раствором DNase Stop (Invitrogen), осадок клеток цианобактерий трехкратно промывали стерильной водой.Последнее лечение было направлено на разрушение клеток цианобактерий и, наконец, на извлечение ДНК цианобактерий, и было основано на протоколе, описанном Тиллеттом и Нейланом (2000), который был слегка изменен, добавив этап механического разрушения клеток (стеклянные шарики и бисер) перед экстракцией ДНК. .
Первая обработка была применена к здоровым культурам в середине активного роста и была основана на ранее опубликованном протоколе для клеток, богатых сахаром (Sharma et al., 2002), с несколькими модификациями, подробно описанными на дополнительном рисунке S1. Для обеспечения деградации оставшейся ДНК нецианобактерий осадок клеток цианобактерий, полученный после первой обработки, ресуспендировали в 100 мкл стерильной воды, содержащей четыре единицы DN азы I (Invitrogen), и инкубировали при 37°C в течение 1 час Ферментативную реакцию останавливали при 65°С раствором DNase Stop (Invitrogen), осадок клеток цианобактерий трехкратно промывали стерильной водой.Последнее лечение было направлено на разрушение клеток цианобактерий и, наконец, на извлечение ДНК цианобактерий, и было основано на протоколе, описанном Тиллеттом и Нейланом (2000), который был слегка изменен, добавив этап механического разрушения клеток (стеклянные шарики и бисер) перед экстракцией ДНК. .
Секвенирование генома, сборка и аннотация
Gloeocapsopsis sp. ДНК UTEX B3054 использовали для секвенирования генома с помощью Illumina MiSeq. Данные были проанализированы с использованием конвейера Illumina 1.4.0 для создания файлов fastq. Необработанные последовательности очищали от штрих-кода, качество проверяли с помощью программного обеспечения FastQC (Andrews, 2010), а считывания обрабатывали с помощью Trimmomatic (Bolger et al., 2014). Обрезанные чтения представляли собой de novo , собранных в 25 153 контига общей длиной 18 225 500 п.н. с использованием Spades v.3.9.0 (Банкевич и др., 2012). Контиги размером менее 3000 п.н. были исключены из окончательного пула последовательностей, поскольку их наиболее близкое совпадение по нуклеотидной идентичности не было цианобактериальным, после чего была проведена вторая сборка de novo с использованием только соответствующих обрезанных прочтений из контигов >3000 п.н.В этой сборке все контиги с низким уровнем глубинного секвенирования соответствовали последовательностям ДНК гетеротрофных бактерий и поэтому были исключены.
ДНК UTEX B3054 использовали для секвенирования генома с помощью Illumina MiSeq. Данные были проанализированы с использованием конвейера Illumina 1.4.0 для создания файлов fastq. Необработанные последовательности очищали от штрих-кода, качество проверяли с помощью программного обеспечения FastQC (Andrews, 2010), а считывания обрабатывали с помощью Trimmomatic (Bolger et al., 2014). Обрезанные чтения представляли собой de novo , собранных в 25 153 контига общей длиной 18 225 500 п.н. с использованием Spades v.3.9.0 (Банкевич и др., 2012). Контиги размером менее 3000 п.н. были исключены из окончательного пула последовательностей, поскольку их наиболее близкое совпадение по нуклеотидной идентичности не было цианобактериальным, после чего была проведена вторая сборка de novo с использованием только соответствующих обрезанных прочтений из контигов >3000 п.н.В этой сборке все контиги с низким уровнем глубинного секвенирования соответствовали последовательностям ДНК гетеротрофных бактерий и поэтому были исключены. Для расчета полноты и качества (степени загрязнения) полученного генома использовали инструмент CheckM v1.0.5 (Parks et al., 2015). Ближайшие доступные геномы были проанализированы с помощью Tetra Correlation Search (TCS) и по средней идентичности нуклеотидов (ANI) с использованием инструмента JSpecies (Richter et al., 2015).
Для расчета полноты и качества (степени загрязнения) полученного генома использовали инструмент CheckM v1.0.5 (Parks et al., 2015). Ближайшие доступные геномы были проанализированы с помощью Tetra Correlation Search (TCS) и по средней идентичности нуклеотидов (ANI) с использованием инструмента JSpecies (Richter et al., 2015).
Автоматическая аннотация генома была проведена с помощью программы PROKKA v.1.11 (Seemann, 2014), и оно было отправлено в JGI IMG/ER для аннотации (ID проекта GOLD Analysis: Ga0181813). Данные о последовательности генома и аннотации доступны в базе данных JGI IMG/ER (идентификатор таксона GOLD: 2756170284; проект секвенирования: Gp0208497 и идентификатор проекта анализа: Ga0181813). Дальнейшая ручная аннотация была выполнена для ограниченного набора представляющих интерес генов, а именно тех, которые участвуют в биосинтезе совместимых растворенных веществ сахарозы и трегалозы, а также генов, связанных с оболочкой. Поиск этих генов осуществлялся с использованием результатов автоматического аннотирования в виде запроса, но дополняя их выполнением поиска BLASTP (Altschul et al. , 1997) для гомологии последовательностей с использованием аминокислотных последовательностей охарактеризованных бактериальных ортологов представляющих интерес генов (используемый порог: e — значение < e -10 и битовая оценка > 30). Кроме того, все предполагаемые гены-ортологи, обнаруженные в геноме Gloeocapsopsis , были проанализированы в базе данных InterProScan, чтобы подтвердить наличие специфических функциональных доменов. В случае генов, кодирующих гликозилтрансферазу, классификация на различные семейства на основе последовательностей выполнялась в соответствии с классификацией, доступной на веб-сайте CAZy (Carbohydrate-Active EnZymes) (Coutinho et al., 2003). Наконец, все предполагаемые интересующие гены, обнаруженные в Gloeocapsopsis sp. UTEX B3054 использовали в качестве запроса для двунаправленного поиска BLASTP в отношении родственных цианобактериальных геномов (используемый порог: e — значение <1 e -15 и битовая оценка > 100).
, 1997) для гомологии последовательностей с использованием аминокислотных последовательностей охарактеризованных бактериальных ортологов представляющих интерес генов (используемый порог: e — значение < e -10 и битовая оценка > 30). Кроме того, все предполагаемые гены-ортологи, обнаруженные в геноме Gloeocapsopsis , были проанализированы в базе данных InterProScan, чтобы подтвердить наличие специфических функциональных доменов. В случае генов, кодирующих гликозилтрансферазу, классификация на различные семейства на основе последовательностей выполнялась в соответствии с классификацией, доступной на веб-сайте CAZy (Carbohydrate-Active EnZymes) (Coutinho et al., 2003). Наконец, все предполагаемые интересующие гены, обнаруженные в Gloeocapsopsis sp. UTEX B3054 использовали в качестве запроса для двунаправленного поиска BLASTP в отношении родственных цианобактериальных геномов (используемый порог: e — значение <1 e -15 и битовая оценка > 100). В частности, в случае генов, предположительно связанных с биосинтезом и транспортом трегалозы, автоматическая аннотация идентифицировала несколько предполагаемых ортологов, но с низким уровнем идентичности аминокислотной последовательности (<35%).Таким образом, чтобы подтвердить гомологию белка, на веб-сервере Phyre2 было выполнено дополнительное трехмерное предсказание структуры белка (Kelly et al., 2015) с целью сделать вывод о структурной гомологии предсказанных белков, где определенные функциональные домены отсутствовали и / или когда идентичность последовательности была ниже 35%.
В частности, в случае генов, предположительно связанных с биосинтезом и транспортом трегалозы, автоматическая аннотация идентифицировала несколько предполагаемых ортологов, но с низким уровнем идентичности аминокислотной последовательности (<35%).Таким образом, чтобы подтвердить гомологию белка, на веб-сервере Phyre2 было выполнено дополнительное трехмерное предсказание структуры белка (Kelly et al., 2015) с целью сделать вывод о структурной гомологии предсказанных белков, где определенные функциональные домены отсутствовали и / или когда идентичность последовательности была ниже 35%.
Анализ генома
Древо филогеномных видов было создано путем объединения аминокислотных последовательностей 43 маркерных генов цианобактерий (Parks et al., 2015).Эти последовательности были извлечены из Gloeocapsopsis sp. Геном UTEX B3054 и другие геномы 74 цианобактерий доступны в базе данных NCBI с помощью инструмента CheckM версии 1.0.5 (Parks et al., 2015). В случае дерева ABC sug последовательности также были извлечены из базы данных NCBI и соответствовали 33 различным геномам, включая Gloeocapsopsis sp. УТЕКС B3054. Последовательности выравнивали с помощью программного обеспечения MUSCLE v6.0.98 (Edgar, 2004). Деревья максимального правдоподобия были сгенерированы с помощью IQtree v.1.5.5 с непараметрической поддержкой ультразагрузки 10 000 повторов (Hoang et al., 2017). Для многолокусного дерева, а также для деревьев TreH и SugABC лучшие модели эволюционной замены были выбраны из выравнивания последовательностей с помощью опции Modelfinder, содержащейся в программном обеспечении IQTree.
УТЕКС B3054. Последовательности выравнивали с помощью программного обеспечения MUSCLE v6.0.98 (Edgar, 2004). Деревья максимального правдоподобия были сгенерированы с помощью IQtree v.1.5.5 с непараметрической поддержкой ультразагрузки 10 000 повторов (Hoang et al., 2017). Для многолокусного дерева, а также для деревьев TreH и SugABC лучшие модели эволюционной замены были выбраны из выравнивания последовательностей с помощью опции Modelfinder, содержащейся в программном обеспечении IQTree.
Анализы потенциального горизонтального переноса генов были выполнены с использованием программного обеспечения HGTector (Zhu et al., 2014) с учетом 50% идентичности аминокислот и 80% охвата последовательностей для результатов BLASTP.С помощью веб-сервера AntiSMASH 4.2.0 (Blin et al., 2017) были обнаружены общие пути биосинтеза вторичных метаболитов. Поиск включал ClusterFinder и полногеномный анализ PFAM.
Результаты
Получение очищенной и нерасщепленной ДНК цианобактерий, подходящей для секвенирования генома
Для получения высококачественной ДНК (соотношение 260/280 > 1,8), пригодной для секвенирования генома богатого сахаром Gloeocapsopsis , потребовалось массовое уничтожение гетеротрофных бактерий для получения чистой и клональной культуры, что было выполнено однократным сортировка клеток в сочетании с проточной цитометрией. Однако даже после последующего штрихования и посева на агар культура оставалась не аксенной (не показано). После этого была разработана трехэтапная процедура, направленная на (1) устранение гетеротрофных бактерий и дестабилизацию агрегатов цианобактерий; (2) устранение оставшихся последовательностей гетеротрофных бактерий; и (3) окончательное разрушение клеток цианобактерий для извлечения чистой ДНК цианобактерий (дополнительная фигура S2). Заключительный этап выделения ДНК позволил эффективно разрушить клетки цианобактерий и избирательно удалить остаточные молекулы сахара.Этот протокол привел к образцу недеградированной ДНК с явным преобладанием ДНК цианобактерий, пригодной для секвенирования (дополнительные рисунки S2B, C).
Однако даже после последующего штрихования и посева на агар культура оставалась не аксенной (не показано). После этого была разработана трехэтапная процедура, направленная на (1) устранение гетеротрофных бактерий и дестабилизацию агрегатов цианобактерий; (2) устранение оставшихся последовательностей гетеротрофных бактерий; и (3) окончательное разрушение клеток цианобактерий для извлечения чистой ДНК цианобактерий (дополнительная фигура S2). Заключительный этап выделения ДНК позволил эффективно разрушить клетки цианобактерий и избирательно удалить остаточные молекулы сахара.Этот протокол привел к образцу недеградированной ДНК с явным преобладанием ДНК цианобактерий, пригодной для секвенирования (дополнительные рисунки S2B, C).
Особенности генома и филогения
После секвенирования ДНК этапы биоинформатической фильтрации отбрасывали оставшиеся последовательности, не относящиеся к цианобактериям (что отражено в двух дискретных пиках содержания G+C, при 43 и 68%), что позволило нам дифференциально собрать геном цианобактерий с полнотой 100 и 1,4% загрязнение последовательности, обнаруженное инструментом CheckM. Полученный размер генома составляет 5 478 916 п.н. в 105 контигах в диапазоне от 3 290 до 356 189 п.н. N 50 соответствует 90 437 п.н. с окончательным охватом генома 138,33X. Содержание G+C составляет 42,41%, представлено одним пиком. Размер генома и содержание G+C аналогичны таковым у Gloeocapsa sp. PCC7438 (5,9 Мб и 42,45% соответственно), самая близкая по последовательности цианобактерия к Gloeocapsopsis sp. УТЕКС B3054. В результате автоматической аннотации генома с помощью программного обеспечения PROKKA было получено в общей сложности 5165 кодирующих белок последовательностей (CDS), 40 тРНК, 3 рРНК и 3 массива CRISPR.Среди первых 1767 последовательностей соответствуют гипотетическим белкам. Общие характеристики генома Gloeocapsopsis по сравнению с другими полностью секвенированными геномами цианобактерий показаны в таблице 1.
Полученный размер генома составляет 5 478 916 п.н. в 105 контигах в диапазоне от 3 290 до 356 189 п.н. N 50 соответствует 90 437 п.н. с окончательным охватом генома 138,33X. Содержание G+C составляет 42,41%, представлено одним пиком. Размер генома и содержание G+C аналогичны таковым у Gloeocapsa sp. PCC7438 (5,9 Мб и 42,45% соответственно), самая близкая по последовательности цианобактерия к Gloeocapsopsis sp. УТЕКС B3054. В результате автоматической аннотации генома с помощью программного обеспечения PROKKA было получено в общей сложности 5165 кодирующих белок последовательностей (CDS), 40 тРНК, 3 рРНК и 3 массива CRISPR.Среди первых 1767 последовательностей соответствуют гипотетическим белкам. Общие характеристики генома Gloeocapsopsis по сравнению с другими полностью секвенированными геномами цианобактерий показаны в таблице 1.
Таблица 1. Общие характеристики Gloeocapsopsis sp. UTEX B3054 и другие геномы цианобактерий.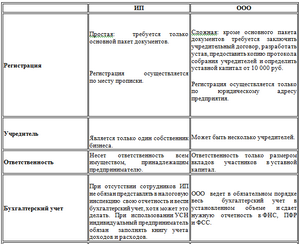
Филогеномный анализ показал, что Gloeocapsopsis sp. UTEX B3054 сформировал отдельную кладу с одноклеточными цианобактериями Gloeocapsa sp.PCC7428 и Chroogloeocystis siderophila NIES-1031 (100% поддержки узла и ~82% ANI) (рис. 1, в сером поле). Семейство Chroococcaceae (порядок Chroococcales), представленное здесь Gloeocapsopsis sp. UTEX B3054 хорошо отделен от одноклеточного Chroococcidiopsis Thermalis PCC7203, но в основном связан с ним (97% поддержки узлов и 70,40% ANI) (рис. 1). Наше филогенетическое дерево подтверждает многоклеточное происхождение этих одноклеточных цианобактерий.
Рис. 1. (A) Просвечивающая электронная микрофотография характерной тетрады одноклеточного Gloeocapsopsis sp. УТЕКС B3054. Обратите внимание на многослойное и обильное внеклеточное полимерное вещество, окружающее клетки. Масштабная линейка: 1 мкм. (B) Филогеномное дерево, созданное методом максимального правдоподобия с использованием 43 маркерных генов цианобактерий из 74 геномов цианобактерий на основе модели эволюционных замен LG+R8. Gloeobacter violaceus PCC7421 использовали в качестве внешней группы.В сером прямоугольнике указано семейство Chroococcaceae (порядок Chroococcales), которое включает Gloeocapsopsis sp. УТЕКС B3054. Эта клада имеет общих предков с Chroococcidiopsis Thermalis PCC7420, который был назначен Komárek et al. (2014) к семейству Chroococcidiopsidaceae (отряд Chroococcidiopsidales). Несколько геномов представителей обеих клад были успешно секвенированы. Значения начальной загрузки из филогенетического анализа отображаются в узлах. В белой рамке представители цианобактерий раздела IV и отдела V.Обе клады относятся к отряду Nostocales.
Gloeobacter violaceus PCC7421 использовали в качестве внешней группы.В сером прямоугольнике указано семейство Chroococcaceae (порядок Chroococcales), которое включает Gloeocapsopsis sp. УТЕКС B3054. Эта клада имеет общих предков с Chroococcidiopsis Thermalis PCC7420, который был назначен Komárek et al. (2014) к семейству Chroococcidiopsidaceae (отряд Chroococcidiopsidales). Несколько геномов представителей обеих клад были успешно секвенированы. Значения начальной загрузки из филогенетического анализа отображаются в узлах. В белой рамке представители цианобактерий раздела IV и отдела V.Обе клады относятся к отряду Nostocales.
Анализ с помощью программного обеспечения HGTector показал, что количество потенциально переносимых по горизонтали CDS составляет около 200, являясь следующими основными донорами бактериального порядка: Bacillales (15%), Ktedonobacterales (7%), Myxococcales (7%) и Rhizobiales (6%). %). Из этих генов 42% обладают неизвестными функциями, 35% связаны с известными метаболическими путями, а 16,5% участвуют в хранении и обработке информации. Гены, связанные с метаболизмом и транспортом вторичных метаболитов, липидов и сахаридов, являются наиболее повторяющимися (дополнительная таблица S1).Кроме того, методом ClusterFinder было идентифицировано 38 кластеров, кодирующих предполагаемые пути вторичных метаболитов. Из них 13 кластеров генов могут быть связаны с биосинтезом сахаридов. Особый интерес представляют восемь кластеров генов, обнаруженных с помощью программного обеспечения antiSMASH, которые кодируют предполагаемые пути биосинтеза бактериоцина (1), терпенов (1), аминогликозид-аминоциклитола (1), поликетидсинтазы (PKS) (1), нерибосомального пептида. синтез (NRPS) (1) и гибриды NRPS-PKS (3). Примечательно, что одна из последних особенно расширена (72.8 т.п.н.), включающий почти 50 генов (Contig00007; расположение 61 046–133 879 н.). В базе данных IMG/JGI не было обнаружено полных гомологов этого кластера генов, что подтверждает его уникальность. В целом эти восемь кластеров генов составляют 4,92% (269 747 п.н.) всего генома Gloeocapsopsis sp.
Гены, связанные с метаболизмом и транспортом вторичных метаболитов, липидов и сахаридов, являются наиболее повторяющимися (дополнительная таблица S1).Кроме того, методом ClusterFinder было идентифицировано 38 кластеров, кодирующих предполагаемые пути вторичных метаболитов. Из них 13 кластеров генов могут быть связаны с биосинтезом сахаридов. Особый интерес представляют восемь кластеров генов, обнаруженных с помощью программного обеспечения antiSMASH, которые кодируют предполагаемые пути биосинтеза бактериоцина (1), терпенов (1), аминогликозид-аминоциклитола (1), поликетидсинтазы (PKS) (1), нерибосомального пептида. синтез (NRPS) (1) и гибриды NRPS-PKS (3). Примечательно, что одна из последних особенно расширена (72.8 т.п.н.), включающий почти 50 генов (Contig00007; расположение 61 046–133 879 н.). В базе данных IMG/JGI не было обнаружено полных гомологов этого кластера генов, что подтверждает его уникальность. В целом эти восемь кластеров генов составляют 4,92% (269 747 п.н.) всего генома Gloeocapsopsis sp. УТЕКС B3054.
УТЕКС B3054.
Совместимые растворенные вещества: биосинтез сахарозы и трегалозы
Два гена, участвующих в биосинтезе сахарозы, обнаружены у Gloeocapsopsis sp. Геном UTEX B3054.Ген фермента сахарозо-6-фосфатсинтазы длиной 2420 п.н. (идентификатор локуса гена: BWI75_00738) был расположен в геномной области, лишенной других генов, связанных с метаболизмом сахара. Его предсказанная аминокислотная последовательность содержит два характерных ферментативных домена ферментов сахарозосинтазы (записи InterProScan: IPR000368 и IPR001296), что указывает на то, что этот белок принадлежит к семейству 1 гликозилтрансфераз (GT). Напротив, 806-пн ген фермента сахарозо-6-фосфатфосфатазы (идентификатор локуса гена: BWI75_01845) был расположен выше кластера генов, вероятно, связанного с метаболизмом сахара (идентификаторы локуса гена: от BWI75_01850 до BWI75_01856), содержащего три гена гликозилтрансферазы в тандеме. .Его предсказанная аминокислотная последовательность содержит домен сахарозофосфатазы (запись InterProScan: IPR006380) и HAD-подобный домен галоиддегидрогеназы (запись InterProScan: IPR023214).
В свою очередь, были идентифицированы два предполагаемых гена, кодирующих белок TreS трегалозосинтазы (идентификаторы локусов генов: BWI75_02890 и BWI75_05160), которые более чем на 50% аминокислотно идентичны с TreS устойчивой к высыханию цианобактерии Leptolyngbya ohadii (Murik et al. ., 2017). Со 100% достоверностью трехмерные прогнозы, сделанные разработчиком структурных моделей Phyre2, показали, что BWI75_02890 и BWI75_05160 имеют 36 и 40% идентичности последовательностей, соответственно, с белком трегалозосинтазы из Mycobacterium smegmatis , одним из наиболее характерных ферментов трегалозосинтазы. в бактериальном мире (Ruhal et al., 2013). Однако у Gloeocapsopsis sp. Геном UTEX B3054.
Ортолог Anabaena sp. Ген трегалазы PCC7120 ( tre H) также был идентифицирован в геноме Gloeocapsopsis (идентификатор локуса гена: BWI75_03170), обладая идентичностью аминокислот на 63,87% (параметры выравнивания: e — значение 0. 0; 584 балла). Со 100% достоверностью трехмерные прогнозы специалиста по моделированию структур Phyre2 показали, что BWI75_03170 имеет 37% идентичности последовательности с ферментом, охарактеризованным как нейтральная трегалаза из дрожжей Saccharomyces cerevisiae , Nth2. Филогенетическая реконструкция показала раннее расхождение белков трегалазы из семейства Chroococcaceae по сравнению с родственными цианобактериями, такими как Nostocales (рис. 2). Интересно, что только в семействе Chroococcaceae ген treH вставлен в консервативный кластер генов, включающий транспортную систему ABC sug ABC (идентификаторы локусов генов: BWI75_03173; BWI75_03172; BWI75_03171 соответственно) и сахар-связывающий белок, расположенный выше . treH (идентификатор локуса гена: BWI75_03174), а также гомосеринкиназу (идентификатор локуса гена: BWI75_03175) (рис. 3).
0; 584 балла). Со 100% достоверностью трехмерные прогнозы специалиста по моделированию структур Phyre2 показали, что BWI75_03170 имеет 37% идентичности последовательности с ферментом, охарактеризованным как нейтральная трегалаза из дрожжей Saccharomyces cerevisiae , Nth2. Филогенетическая реконструкция показала раннее расхождение белков трегалазы из семейства Chroococcaceae по сравнению с родственными цианобактериями, такими как Nostocales (рис. 2). Интересно, что только в семействе Chroococcaceae ген treH вставлен в консервативный кластер генов, включающий транспортную систему ABC sug ABC (идентификаторы локусов генов: BWI75_03173; BWI75_03172; BWI75_03171 соответственно) и сахар-связывающий белок, расположенный выше . treH (идентификатор локуса гена: BWI75_03174), а также гомосеринкиназу (идентификатор локуса гена: BWI75_03175) (рис. 3).
Рисунок 2. Филогения цианобактериальной трегалазы. Дерево максимального правдоподобия было создано с помощью программного обеспечения IQTree на основе модели эволюционной замены LG+I+G4. Белки трегалазы из семейства Chroococcaceae (серая рамка) демонстрируют раннюю дивергенцию по сравнению с Nostocales и другими цианобактериями. Числа в узлах указывают значения начальной загрузки из филогенетического анализа.
Белки трегалазы из семейства Chroococcaceae (серая рамка) демонстрируют раннюю дивергенцию по сравнению с Nostocales и другими цианобактериями. Числа в узлах указывают значения начальной загрузки из филогенетического анализа.
Рисунок 3. Организация локуса sug A– sug B– sug C у цианобактерий.Дерево максимального правдоподобия было создано с помощью программного обеспечения IQTree на основе модели эволюционной замены LG+I+G4. Только цианобактерии из семейства Chroococcaceae имеют tre H, фланкирующие 3′-конец генов sug A– sug B– sug C. Отметим также, что в этом семействе гомосеринкиназа и белок, связывающий транспорт ABC, расположены на 5′-конце этого кластера генов.
Гены, связанные с оболочкой: биосинтез и экспорт ЭПС
Был проведен геномный скрининг для идентификации всех предполагаемых белков гликозилтрансферазы (GT), кодируемых Gloeocapsopsis sp.UTEX B3054 и родственные цианобактерии. Тщательный поиск показал, что Gloeocapsopsis sp. UTEX B3054 обладает по крайней мере 129 генами, кодирующими GTs (Таблица 1), столько же, сколько у его ближайшего эволюционного родственника Gloeocapsa sp. ПКС7428. C. Thermalis PCC7203 , Gloeocapsa sp. PCC7428 и Gloeocapsopsis sp. UTEX B3054 обладает самым высоким соотношением GTs/CDS (табл. 1) по сравнению с Anabaena sp. PCC7120, Nostoc punctiforme и Synechocystis sp.ПКС6803. Gloeocapsopsis sp. UTEX B3054 и Gloeocapsa sp. PCC7428 представляет высший уровень разнообразия GT с 30 различными типами. Synechocystis зр. PCC6803 содержит наименьшее разнообразие GT, кодируя только 21 тип. Результаты показывают, что большинство предсказанных цианобактериальных GT организованы в виде функциональных модулей с консервативными ферментативными мотивами, структурированными в тандеме (дополнительные таблицы S2-S6), и несколькими различными предсказанными доменами, объединенными в большой пул предполагаемых белков GT. Большинство GT, обнаруженных в Gloeocapsopsis sp. Геномы UTEX B3054 и других цианобактерий (более 70%) принадлежат к семействам GT1 и GT2 (дополнительные таблицы S2–S6). Было обнаружено, что многие гены GT встроены в геномные кластеры, связанные с метаболизмом сахара, например, в контиге PWI75_000047 (рис. 4), который содержит 10 генов GT и дополнительные гены, связанные с метаболизмом сахара, чьи ортологи в Anabaena sp. PCC7120 активируются при высыхании (Yoshimura et al., 2007).
Тщательный поиск показал, что Gloeocapsopsis sp. UTEX B3054 обладает по крайней мере 129 генами, кодирующими GTs (Таблица 1), столько же, сколько у его ближайшего эволюционного родственника Gloeocapsa sp. ПКС7428. C. Thermalis PCC7203 , Gloeocapsa sp. PCC7428 и Gloeocapsopsis sp. UTEX B3054 обладает самым высоким соотношением GTs/CDS (табл. 1) по сравнению с Anabaena sp. PCC7120, Nostoc punctiforme и Synechocystis sp.ПКС6803. Gloeocapsopsis sp. UTEX B3054 и Gloeocapsa sp. PCC7428 представляет высший уровень разнообразия GT с 30 различными типами. Synechocystis зр. PCC6803 содержит наименьшее разнообразие GT, кодируя только 21 тип. Результаты показывают, что большинство предсказанных цианобактериальных GT организованы в виде функциональных модулей с консервативными ферментативными мотивами, структурированными в тандеме (дополнительные таблицы S2-S6), и несколькими различными предсказанными доменами, объединенными в большой пул предполагаемых белков GT. Большинство GT, обнаруженных в Gloeocapsopsis sp. Геномы UTEX B3054 и других цианобактерий (более 70%) принадлежат к семействам GT1 и GT2 (дополнительные таблицы S2–S6). Было обнаружено, что многие гены GT встроены в геномные кластеры, связанные с метаболизмом сахара, например, в контиге PWI75_000047 (рис. 4), который содержит 10 генов GT и дополнительные гены, связанные с метаболизмом сахара, чьи ортологи в Anabaena sp. PCC7120 активируются при высыхании (Yoshimura et al., 2007).
Рис. 4.Кластер генов, вероятно, связан с механизмами устойчивости к высыханию. Этот кластер генов содержит 10 генов, кодирующих гликозилтрансферазы, ортологи которых в Anabaena продемонстрировали сверхэкспрессию в условиях высушивания (Yoshimura et al., 2007). Локус гена: от PWI75_04273 до PWI75_04293.
Заключительный этап биосинтеза внеклеточных полисахаридов включает сборку и транслокацию полисахаридных цепей во внеклеточное пространство. Этот этап может выполняться тремя основными путями: зависимым от Wzy, зависимым от ABC-транспортера и зависимым от синтазы (Pereira et al., 2009, 2015; Кер и Диттманн, 2015). В Gloeocapsopsis sp. Были обнаружены UTEX B3054, ортологи генов, ассоциированных с тремя путями экспорта полисахаридов (табл. 2). Всего 223 гена, представляющие 6,89% Gloeocapsopsis sp. Геном UTEX B3054 включен в кластер ортологичных генов (COG) категории «Биогенез клеточной стенки/мембраны/оболочки» (не показано).
Этот этап может выполняться тремя основными путями: зависимым от Wzy, зависимым от ABC-транспортера и зависимым от синтазы (Pereira et al., 2009, 2015; Кер и Диттманн, 2015). В Gloeocapsopsis sp. Были обнаружены UTEX B3054, ортологи генов, ассоциированных с тремя путями экспорта полисахаридов (табл. 2). Всего 223 гена, представляющие 6,89% Gloeocapsopsis sp. Геном UTEX B3054 включен в кластер ортологичных генов (COG) категории «Биогенез клеточной стенки/мембраны/оболочки» (не показано).
Таблица 2. Гены, связанные с путями экспорта ЭПС у Gloeocapsopsis sp.УТЕКС B3054.
Обсуждение
Как и у многих культивируемых цианобактерий, острота Gloeocapsopsis оказалась одной из главных проблем. Первоначальные попытки в нашей лаборатории секвенировать геном Gloeocapsopsis подтвердили персистенцию гетеротрофных бактерий в культуре, что помешало эффективному секвенированию генома цианобактерий. Таким образом, в первой попытке секвенировать обогащение AAB1 с использованием технологии пиросеквенирования 454, всего было получено 19,8 Мб, поразительно 95. 5% соответствовали гетеротрофным бактериям, принадлежащим к 12 разным родам. В настоящем исследовании сортировка отдельных клеток позволила нам преодолеть эту ситуацию, восстановив менее загрязненную клональную культуру, по крайней мере, до приемлемой степени.
5% соответствовали гетеротрофным бактериям, принадлежащим к 12 разным родам. В настоящем исследовании сортировка отдельных клеток позволила нам преодолеть эту ситуацию, восстановив менее загрязненную клональную культуру, по крайней мере, до приемлемой степени.
Наши результаты продемонстрировали, что все технические усилия, предпринятые в этом исследовании для эффективного и успешного разрушения внеклеточной полисахаридной структуры, к которой прикреплены гетеротрофные бактерии, укрепили представление о том, что Gloeocapsopsis sp.Оболочка UTEX B3054 богата сахаром и впечатляюще трудно поддается разрушению: клетки невосприимчивы к механическим, физическим, химическим и биологическим методам разрушения. Таким образом, в молекулярных исследованиях, направленных на раскрытие микробного разнообразия и метаболических свойств микробов-экстремофилов, процедура выделения ДНК должна вызывать серьезную озабоченность (Lever et al., 2015).
Реализация трехэтапной процедуры позволила дифференцированно выделить ДНК цианобактерий, получив качественный материал для секвенирования. Этот окончательный протокол избегал совместного осаждения сахаров и загрязняющей ДНК с желаемой ДНК цианобактерий, которые, вероятно, мешали амплификации ДНК с помощью ПЦР, эффективности клонирования и дальнейшему секвенированию и сборке генома (Angeloni and Potts, 1987; Billi et al., 1998; Fiore et al., 2000; Tillett and Neilan, 2000; Chrismas et al., 2016). По нашим оценкам, этот протокол может быть адаптирован для других труднолизируемых и богатых сахаром неаксеновых цианобактерий.
Этот окончательный протокол избегал совместного осаждения сахаров и загрязняющей ДНК с желаемой ДНК цианобактерий, которые, вероятно, мешали амплификации ДНК с помощью ПЦР, эффективности клонирования и дальнейшему секвенированию и сборке генома (Angeloni and Potts, 1987; Billi et al., 1998; Fiore et al., 2000; Tillett and Neilan, 2000; Chrismas et al., 2016). По нашим оценкам, этот протокол может быть адаптирован для других труднолизируемых и богатых сахаром неаксеновых цианобактерий.
Следует отметить, что одновременно с представлением этого отчета другая группа, работающая с исходной зараженной культурой Gloeocapsopsis , опубликовала ее геном (Puente-Sanchez et al., 2018). Несмотря на то, что наши протоколы выделения клеток и извлечения ДНК отнимали много времени, они помогли нам получить более точные геномные данные, т. е. мы получили меньше контигов (105 против 137), а наш самый короткий контиг имеет 3290 п.н. (против только 137 п.н.), в то время как самый длинный содержит 356 189 п. н. (против 250 842 п.н.). Параметр N 50 нашей сборки оказался выше (90 347 против 73 596). Более того, благодаря тому, что геномы цианобактерий характеризуются протяженными повторяющимися участками (Mazel et al., 1990; Asayama et al., 1996; Elhai et al., 2008), индивидуальный протокол выделения ДНК, способный гарантировать, что высококачественные молекулы ДНК имеют решающее значение для облегчения обработки биоинформатики.
н. (против 250 842 п.н.). Параметр N 50 нашей сборки оказался выше (90 347 против 73 596). Более того, благодаря тому, что геномы цианобактерий характеризуются протяженными повторяющимися участками (Mazel et al., 1990; Asayama et al., 1996; Elhai et al., 2008), индивидуальный протокол выделения ДНК, способный гарантировать, что высококачественные молекулы ДНК имеют решающее значение для облегчения обработки биоинформатики.
Уже известно, что цианобактерии накапливают сахарозу и трегалозу не только в цитоплазме, но и во внеклеточном матриксе как механизм устойчивости к высыханию (Sakamoto et al., 2009, 2011; Yoshida, Sakamoto, 2009; Azúa-Bustos et al. , 2014; Мурик и др., 2017). Геномный анализ Gloeocapsopsis sp.UTEX B3054 выявил наличие генов биосинтеза сахарозы и трегалозы. Две формы фермента сахарозофосфатсинтазы (SPS) были обнаружены у цианобактерий (Blank, 2013), и Gloeocapsopsis кодирует только ортолог SPS, родственного нитчатым, который обладает характерным для гликозилтрансферазы единственным функциональным доменом.
Наш геномный анализ выявил наличие только одного набора генов, связанных с биосинтезом трегалозы (гены трегалозсинтазы), и отсутствие генов tre Y/ tre Z. Анализ In silico предсказал, что трегалозосинтаза, вероятно, опосредует биосинтез трегалозы в этой цианобактерии, чьи гомологи treS были транскрипционно активированы в устойчивых к высыханию L. ohadii , подвергнутых моделируемым условиям высыхания (Murik et al., 2017) . В обоих Anabaena sp. PCC7120 и N. punctiforme , концентрация трегалозы регулируется TreH (Yoshida and Sakamoto, 2009), ферментом трегалазой, который кодируется геном, расположенным в кластере генов с мальтоолигозилтрегалозгидролазой ( tre Y) и мальтоолигозилтрегалозсинтазой. ( tre Z) генов (Higo et al., 2006; Йошида и Сакамото, 2009 г.). Однако только в геномах Chroococcaceae ген treH был обнаружен рядом с предполагаемым кластером генов ABC-переносчика сахара, что указывает на существование консервативного механизма регуляции трегалозы, специфичного для этого семейства.
Транспортер ABC, связанный с геном tre H Gloeocapsopsis sp. UTEX B3054 обладает консервативным расположением генома и аминокислотной идентичностью с функционально описанным переносчиком LipqY- Sug ABC Mycobacterium tuberculosis (27.00, 37,13, 38,70 и 42,93% соответственно). У M.tuberculosis этот транспортер обладает высокой специфичностью к трегалозе и в сочетании с его периплазматическим сахаросвязывающим белком работает как эффективная система ретроградной рециркуляции дисахарида, активно участвуя в поглощении трегалозы и биосинтезе внеклеточной оболочки (Kalscheuer et al., 2010). Таким образом, мы предполагаем, что цианобактерии из семейства Chroococcaceae обладают новой системой ретроградной рециркуляции для регулирования концентрации трегалозы, которая может играть решающую роль в сохранении живых клеток во время экстремального высыхания.
В самом недавнем геномном исследовании биосинтеза EPS (Pereira et al., 2015) преднамеренно игнорировалось изучение GTs, несмотря на то, что считается, что они играют центральную роль в бактериальном биосинтезе полисахаридов (Schmid et al. , 2016). Эти ферменты связаны с биосинтезом оболочки, включая ЭПС, липополисахарид и пептидогликан, а также с гликозилированием мембранных липидов и биосинтезом вторичных метаболитов среди других процессов (Campbell et al., 1997; Коутиньо и др., 2003).
, 2016). Эти ферменты связаны с биосинтезом оболочки, включая ЭПС, липополисахарид и пептидогликан, а также с гликозилированием мембранных липидов и биосинтезом вторичных метаболитов среди других процессов (Campbell et al., 1997; Коутиньо и др., 2003).
Полногеномный анализ GTs, проведенный в этой работе, представляет собой первую попытку предсказать весь и разнообразный пул GTs в геномах цианобактерий. Вся классификация GT пока неполная и может быть улучшена по мере того, как будет охарактеризовано больше семейств GT. Уже доступная информация о связанных с оболочкой GTs у цианобактерий сокращена и фрагментирована и, возможно, упускает из виду огромное разнообразие предполагаемых GTs из-за их большого количества и сложности в природе (Yang et al., 2007). Процент от общего количества кодируемых белков, которые могут быть связаны исключительно с синтезом гликозидных связей в Gloeocapsopsis , оказался выше, чем от 1 до 2% от общего количества кодируемых белков, оцененных для других геномов, будь то архей, бактерий или эукариот (табл. 1; Ларсон и др., 2008). Кроме того, Gloeocapsopsis sp. UTEX B3054 кодирует гены-ортологи трех путей, уже описанных у цианобактерий для транспорта и экспорта ЭПС (Pereira et al., 2015), предполагая, что эта цианобактерия может обладать целым молекулярным механизмом для транслокации и экспорта полисахаридов во внеклеточное пространство.
1; Ларсон и др., 2008). Кроме того, Gloeocapsopsis sp. UTEX B3054 кодирует гены-ортологи трех путей, уже описанных у цианобактерий для транспорта и экспорта ЭПС (Pereira et al., 2015), предполагая, что эта цианобактерия может обладать целым молекулярным механизмом для транслокации и экспорта полисахаридов во внеклеточное пространство.
Несколько идентифицированных GT были вставлены в кластеры генов, включающие другие гены, связанные с биосинтезом сахаров, а также несколько кластеров генов, связанных с биосинтезом сахаридов, связанных со вторичными метаболитами. Например, кластер генов, богатый гликозилтрансферазой, обнаруженный в контиге BWI75_000047, оказывается привлекательным кластером для функциональных исследований механизмов толерантности к высыханию.Этот кластер содержит гены, соответствующие ортологи которых активируются у Anabaena sp. PCC7120 при высыхании (Yoshimura et al., 2007). Более того, Gloeocapsopsis sp. UTEX B3054 содержит ортолог сигма-фактора РНК-полимеразы SigJ (идентификатор локуса гена: BWI75_01223), описанного как ключевой регулятор устойчивости к высыханию у Anabaena sp. PCC7120 (68,58% идентичности последовательности) (Yoshimura et al., 2007). Самый длинный кластер генов (Contig00007; местоположение 61 046–133 879 н.), обнаруженный с помощью программного обеспечения antiSMASH, также оказался очень интересной мишенью для дальнейших функциональных исследований: гибридный кластер PKS-сахарид-NRPS, состоящий из 72.8 кб уникальной генетической информации, которая потенциально может представлять собой новый механизм, связанный с высокой устойчивостью этой цианобактерии к экстремальным условиям окружающей среды.
PCC7120 (68,58% идентичности последовательности) (Yoshimura et al., 2007). Самый длинный кластер генов (Contig00007; местоположение 61 046–133 879 н.), обнаруженный с помощью программного обеспечения antiSMASH, также оказался очень интересной мишенью для дальнейших функциональных исследований: гибридный кластер PKS-сахарид-NRPS, состоящий из 72.8 кб уникальной генетической информации, которая потенциально может представлять собой новый механизм, связанный с высокой устойчивостью этой цианобактерии к экстремальным условиям окружающей среды.
Заключение
Сочетание сортировки отдельных клеток и индивидуального протокола многоэтапной экстракции ДНК обеспечило секвенирование Gloeocapsopsis sp. Геном UTEX B3054. Технические трудности, возникшие при извлечении нуклеиновых кислот, подтвердили сложность внеклеточного матрикса и огромное содержание сахара в этом штамме.Клетки продемонстрировали низкую устойчивость к широкому спектру разрушающих методов, что подчеркивает выдающиеся физико-химические свойства их защитной оболочки.
Анализ in silico выявил генетический потенциал для борьбы с дефицитом воды, характерный для Gloeocapsopsis и его родственников. Основные усилия должны быть сосредоточены на расшифровке роли сахаров во время высыхания и, в частности, функциональной роли, которую трегалаза, связанная с переносчиком ABC, может играть в контроле концентраций трегалозы как на внутриклеточном, так и на внеклеточном уровнях.Более того, разнообразие GTs, обнаруженное в этой работе, предполагает, что молекулярная сложность полисахаридной матрицы потенциально может быть огромной.
Мы надеемся, что вся геномная структура, представленная в этой работе, поможет распутать состав сахаров и структурное устройство внеклеточного матрикса цианобактерий, чьи реологические свойства, по-видимому, имеют решающее значение для удержания скудной воды в Атакаме.
Вклад авторов
CU, RV и BD задумали исследование.CU и LS провели экспериментальную работу и протокол выделения ДНК. CU извлек геномную ДНК для секвенирования под руководством MV, RV и BD. BD запланировала секвенирование генома. MP провел секвенирование генома. CU и JA провели геномное исследование под руководством BD. CU, JA, MV, RV и BD проанализировали данные. CU, RV и BD написали рукопись. Все авторы прочитали и прокомментировали черновик рукописи.
CU извлек геномную ДНК для секвенирования под руководством MV, RV и BD. BD запланировала секвенирование генома. MP провел секвенирование генома. CU и JA провели геномное исследование под руководством BD. CU, JA, MV, RV и BD проанализировали данные. CU, RV и BD написали рукопись. Все авторы прочитали и прокомментировали черновик рукописи.
Финансирование
Эта работа была частично поддержана грантами FONDECYT (1110597, 1150171 и 1161232) и Института фундаментальной и прикладной биологии тысячелетия (MIFAB).CU финансировался за счет докторской стипендии CONICYT, а также Beca Gastos Operationales CONICYT 21110394. Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы поблагодарить Германа Марчандона за его помощь и руководство в процессе сборки генома.CU хотел бы особо поблагодарить доктора Ger van den Engh за помощь в получении культуры униалгала с помощью проточной цитометрии в сочетании с сортировкой клеток.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00950/full#supplementary-material
.Каталожные номера
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST и PSI-BLAST: новое поколение программ поиска белковых баз данных. Рез. нуклеиновых кислот. 25, 3389–3402.
Реферат PubMed | Академия Google
Ангелони, С.В., и Поттс, М. (1987). Очистка полисом из устойчивой к лизоциму устойчивой к высыханию цианобактерии. J. Microbiol. Методы 6, 61–69. дои: 10.1016/0167-7012(87)—
Полнотекстовая перекрестная ссылка | Академия Google
Асаяма М., Кабасава М., Такахаши И., Аида Т. и Шираи М. (1996). Высоко повторяющиеся последовательности и характеристики геномной ДНК у одноклеточных штаммов цианобактерий. FEMS микробиол. лат. 137, 175–181. дои: 10.1016/0378-1097(96)00050-X
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Асуа-Бустос, А., Суньига, Дж., Аренас-Фахардо, К., Орельяна, М., Салас, Л., и Викунья, Р. (2014). Gloeocapsopsis AAB1, чрезвычайно устойчивая к высыханию цианобактерия, выделенная из пустыни Атакама. Экстремофилы 18, 61–74. doi: 10.1007/s00792-013-0592-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бал, Дж., Lau, M.C.Y., Smith, G.J.D., Vijaykrishna, D., Cary, S.C., Lacap, D.C., et al. (2011). Древнее происхождение определяет глобальную биогеографию цианобактерий жарких и холодных пустынь. Нац. коммун. 2, 163–166. doi: 10.1038/ncomms1167
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Банкевич А., Нурк С., Антипов Д., Гуревич А. А., Дворкин М., Куликов А. С. и др. (2012). SPAdes: новый алгоритм сборки генома и его приложения для секвенирования отдельных клеток. Дж. Вычисл. биол. 19, 455–477. doi: 10.1089/cmb.2012.0021
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Билли Д., Грилли Кайола М., Паолоцци Л. и Гелардини П. (1998). Метод выделения ДНК из пустынной цианобактерии Chroococcidiopsis и его применение для идентификации ftsZ. Заяв. Окружающая среда. микробиол. 64, 4053–4056.
Реферат PubMed | Академия Google
Блин, К., Вольф, Т., Шевретт, М.G., Lu, X., Schwalen, C.J., Kautsar, S.A., et al. (2017). AntiSMASH 4.0 — улучшения в предсказании химии и идентификации границ кластера генов. Рез. нуклеиновых кислот. 45, W36–W41. doi: 10.1093/nar/gkx319
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кэмпбелл, Дж. А., Дэвис, Г. Дж., Булоне, В., и Хенриссат, Б. (1997). Классификация нуклеотид-дифосфо-сахар-гликозилтрансфераз, основанная на сходстве аминокислотных последовательностей. Биохим.Дж. 326, 929–939. дои: 10.1042/bj2800309
Полнотекстовая перекрестная ссылка | Академия Google
Chan, Y., Lacap, D.C., Lau, M.C.Y., Ha, K.Y., Warren-Rhodes, K.A., Cockell, C.S., et al. (2012). Гиполитические микробные сообщества: между молотом и наковальней. Окружающая среда. микробиол. 14, 2272–2282. doi: 10.1111/j.1462-2920.2012.02821.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Крисмас, Н. А. М., Баркер, Г., Анесио, А. М., и Санчес-Баракальдо, П.(2016). Геномные механизмы холодоустойчивости и продукции экзополисахаридов у арктической цианобактерии Phormidesmis Priestleli BC1401. BMC Genomics 17:533. doi: 10.1186/s12864-016-2846-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Колика Г., Ли Х., Росси Ф., Ли Д., Лю Ю. и Де Филиппис Р. (2014). Экзополисахариды, секретируемые микробами, влияют на гидрологическое поведение индуцированных биологических почвенных корок в песчаных почвах пустыни. Почвенный биол.Биохим. 68, 62–70. doi: 10.1016/j.soilbio.2013.09.017
Полнотекстовая перекрестная ссылка | Академия Google
Коутиньо П.М., Делери Э., Дэвис Г.Дж. и Хенриссат Б. (2003). Развивающаяся иерархическая семейная классификация гликозилтрансфераз. Дж. Мол. биол. 328, 307–317. doi: 10.1016/S0022-2836(03)00307-303
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коуэн Д. А., Махаланян Т. П., Деннис П. Г. и Хопкинс Д.В. (2014). Микробная экология и биогеохимия почв континентальной Антарктики. Перед. микробиол. 5:154. doi: 10.3389/fmicb.2014.00154
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Crits-Christoph, A., Robinson, C.K., Ma, B., Ravel, J., Wierzchos, J., Ascaso, C., et al. (2016). Филогенетическая и функциональная субстратная специфичность эндолитных микробных сообществ в гипераридных условиях. Перед. микробиол. 7:301. doi: 10.3389/fmicb.2016.00301
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Давила, А.Ф., Хоуз, И., Арайя, Дж.Г., Гелсингер, Д.Р., Ди Руджеро, Дж., Аскасо, К., и др. (2015). Метаболизм in situ в галитовых эндолитных микробных сообществах гипераридной пустыни Атакама. Перед. микробиол. 6:1035. doi: 10.3389/fmicb.2015.01035
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эльхай, Дж., Като, М., Казинс, С., Линдблад, П., и Коста, Дж.Л. (2008). Очень маленькие подвижные повторяющиеся элементы в геномах цианобактерий. Рез. генома. 18, 1484–1499. doi: 10.1101/gr.074336.107
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фиоре М.Ф., Мун Д.Х., Цай С.М., Ли Х. и Треворс Дж.Т. (2000). Выделение минипрепаратов ДНК из одноклеточных и нитчатых цианобактерий. J. Microbiol. Методы 39, 159–169. doi: 10.1016/S0167-7012(99)00110-114
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грилли Кайола, М., Билли Д. и Фридманн Э.И. (1996). Влияние высушивания на оболочки цианобактерий Chroococcidiopsis sp. евро. Дж. Фикол. 31, 37–41.
Академия Google
Grilli Caiola, M., Ocampo-Friedmann, R., and Friedmann, E.I. (1993). Цитология длительного высыхания пустынной цианобактерии Chroococcidiopsis (Chroococcales). Phycologia 32, 315–322.
Реферат PubMed | Академия Google
Хартли, Эй Джей, Чонг, Г., Хьюстон, Дж., и Мазер, А. (2005). 150 миллионов лет стабильности климата: свидетельство пустыни Атакама, север Чили. Дж. Геол. соц. Лонд. 162, 421–424.
Академия Google
Гершковитц, Н., Орен, А., и Коэн, Ю. (1991). Накопление трегалозы и сахарозы у цианобактерий при матриксном водном стрессе. Заяв. Окружающая среда. микробиол. 57, 645–648.
Реферат PubMed | Академия Google
Хиго, А., Като, Х., Омори, К., Икеучи, М., и Омори, М. (2006). Роль кластера генов метаболизма трегалозы в устойчивости к обезвоживанию нитчатых цианобактерий Anabaena sp. PCC 7120. Микробиология 152, 979–987. doi: 10.1099/микрофон 0.28583-28580
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хилл, Д. Р., Кинан, Т. В., Хелм, Р. Ф., Поттс, М., Кроу, Л. М., и Кроу, Дж. Х. (1997). Внеклеточный полисахарид Nostoc commune (цианобактерии) ингибирует слияние мембранных везикул при высыхании. Дж. Заявл. Фикол. 9, 237–248.
Академия Google
Хилл, Д. Р., Пит, А., и Поттс, М. (1994). Биохимия и структура гликана, секретируемого устойчивой к высыханию коммуной Nostoc (цианобактерии). Протоплазма 182, 126–148. дои: 10.1007/BF01403474
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хоанг, Д. Т., Черномор, О., фон Хазелер, А., Минь, Б. К., и Ле, С. В. (2017). UFBoot2: улучшение приближения сверхбыстрой начальной загрузки. Мол. биол. Эвол. 35, 518–522. doi: 10.1093/molbev/msx281
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хьюстон, Дж., и Хартли, А. (2003). Дождевая тень западного склона центральных Анд и ее потенциальный вклад в возникновение гиперзасушливости пустыни Атакама. Междунар. Дж. Климатол. 23, 1453–1464.
Академия Google
Kalscheuer, R., Weinrick, B., Veeraraghavan, U., Besra, G.S., and Jacobs, W.R. (2010). Перерабатывающий трегалозу переносчик ABC LpqY-SugA-SugB-SugC необходим для вирулентности Mycobacterium tuberculosis. Проц. Натл. акад. науч. США 107, 21761–21766. doi: 10.1073/pnas.1014642108
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Келли, Л. А., Мезулис, С., Йейтс, К., Васс, М., и Штернберг, М. (2015). Веб-портал Phyre2 для моделирования, прогнозирования и анализа белков. Нац. протокол 10, 845–858. doi: 10.1038/nprot.2015-2053
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ноулз, Э. Дж., и Кастенхольц, Р.В. (2008). Влияние экзогенных внеклеточных полисахаридов на устойчивость к высыханию и замораживанию фототрофных микроорганизмов, населяющих скалы. FEMS микробиол. Экол. 66, 261–270. doi: 10.1111/j.1574-6941.2008.00568.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Комарек Дж., Каштовский Дж., Мареш Дж. и Йохансен Дж. Р. (2014). Таксономическая классификация цианопрокариот (роды цианобактерий) 2014 г. с использованием полифазного подхода. Преслия 86, 295–335.
Академия Google
Лайрсон, Л.Л., Хенриссат, Б., Дэвис, Г.Дж., и Уизерс, С.Г. (2008). Гликозилтрансферазы: структура, функции и механизмы. год. Преподобный Биохим. 77, 521–555. doi: 10.1146/annurev.biochem.76.061005.092322
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Левер, М. А., Торти, А., Эйкенбуш, П., Мишо, А. Б., Шантл-Темкив, Т., и Йоргенсен, Б. Б. (2015). Модульный метод выделения ДНК и РНК и разделения пулов ДНК из различных типов образцов окружающей среды. Перед. микробиол. 6:476. doi: 10.3389/fmicb.2015.00476
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Магер, Д.М., и Томас, А.Д. (2011). Внеклеточные полисахариды из цианобактериальных почвенных корок: обзор их роли в почвенных процессах засушливых земель. J. Засушливая среда. 75, 91–97. doi: 10.1016/j.jaridenv.2010.10.001
Полнотекстовая перекрестная ссылка | Академия Google
Мазель, Д., Хумар, Дж., Касте, А.М., и Тандо де Марсак, Н.(1990). Высоко повторяющиеся последовательности ДНК в геномах цианобактерий. J. Бактериол. 172, 2755–2761.
Реферат PubMed | Академия Google
Маккей, К.П., Фридманн, Э.И., Гомес-Сильва, Б., Касерес-Вильянуэва, Л., и Андерсен, Д.Т. (2003). Температурно-влажностные условия для жизни в крайне засушливом районе пустыни Атакама: четыре года наблюдений, включая Эль-Ниньо 1997–1998 гг. Астробиология 3, 393–406.
Реферат PubMed | Академия Google
Мурик О., Oren, N., Shotland, Y., Raanan, H., Treves, H., Kedem, I., et al. (2017). Что отличает цианобактерии, способные к возрождению после высыхания, от неспособных: геномный аспект. Окружающая среда. микробиол. 19, 535–550. дои: 10.1111/1462-2920.13486
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Наварро-Гонсалес Р., Рейни Ф. А., Молина П., Багалей Д., Холлен Б. Дж., де ла Роса Дж. и др. (2003). Марсоподобные почвы в пустыне Атакама, Чили, и сухой предел микробной жизни. Наука 302, 1018–1021. doi: 10.1126/наука.1089143
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паркс, Д. Х., Имельфорт, М., Скеннертон, К. Т., Хугенхольц, П., и Тайсон, Г. В. (2015). CheckM: оценка качества микробных геномов, восстановленных из изолятов, отдельных клеток и метагеномов. Рез. генома. 25, 1043–1055. doi: 10.1101/гр.186072.114
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Перейра, С., Zille, A., Micheletti, E., Moradas-Ferreira, P., de Philippis, R. и Tamagnini, P. (2009). Сложность экзополисахаридов цианобактерий: состав, структура, индуцирующие факторы и предполагаемые гены, участвующие в их биосинтезе и сборке. FEMS микробиол. Ред. 33, 917–941. doi: 10.1111/j.1574-6976.2009.00183.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Перейра, С. Б., Мота, Р., Виейра, К. П., Виейра, Дж., и Таманьини, П. (2015). Общефилумный анализ генов/белков, связанных с последними этапами сборки и экспорта внеклеточных полимерных веществ (ВПС) у цианобактерий. науч. Респ. 5:14835. дои: 10.1038/srep14835
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Pointing, S.B., Chan, Y., Lacap, D.C., Lau, M.C.Y., Jurgens, J.A., и Farrell, R.L. (2009). Высокоспециализированное микробное разнообразие гипераридной полярной пустыни. Проц. Натл. акад. науч. США 106, 19964–19969. doi: 10.1073/pnas.0
4106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пуэнте-Санчес, Ф., Гонсалес-Сильва, К., Парро, В., Тамамес, Дж., и Асуа-Бустос, А. (2018). Проект последовательности генома чрезвычайно устойчивой к высыханию цианобактерии Gloecapsopsis sp. штамм AAB 1. Genome Announce. 5, 1–2. doi: 10.1128/genomeA.00216-18
Реферат PubMed | Полный текст перекрестной ссылки
Рихтер, М., Росселло-Мора, Р., Глекнер, Ф. О., и Пеплис, Дж. (2015). JSpeciesWS: веб-сервер для описания прокариотических видов на основе попарного сравнения геномов. Биоинформатика 32, 929–931. doi: 10.1093/биоинформатика/btv681
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Росси, Ф., и Де Филиппис, Р. (2015). Роль экзополисахаридов цианобактерий в фототрофных биопленках и сложных микробных матах. Жизнь 5, 1218–1238. дои: 10.3390/life5021218
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рухал Р., Катариа Р. и Чоудхури Б. (2013). Тенденции метаболизма бактериальной трегалозы и важные узлы метаболического пути в направлении накопления трегалозы. Микроб. Биотехнолог. 6, 493–502. дои: 10.1111/1751-7915.12029
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сакамото Т., Кумихаши К., Кунита С., Масаура Т., Иноуэ-Сакамото К. и Ямагути М. (2011). Сохраняющая внеклеточный матрикс цианобактерия Nostoc verrucosum накапливает трегалозу, но чувствительна к высыханию. FEMS микробиол. Экол. 77, 385–394. doi: 10.1111/j.1574-6941.2011.01114.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сакамото, Т., Йошида Т., Арима Х., Хатанака Ю., Такани Ю. и Тамару Ю. (2009). Накопление трегалозы в ответ на высыхание и солевой стресс у наземных цианобактерий Коммуна Носток . Фикол. Рез. 57, 66–73. doi: 10.1111/j.1440-1835.2008.00522.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шмид Дж., Хайдер Д., Вендель Н. Дж., Сперл Н. и Зибер В. (2016). Бактериальные гликозилтрансферазы: проблемы и возможности очень разнообразного класса ферментов для создания натуральных продуктов. Перед. микробиол. 7:182. doi: 10.3389/fmicb.2016.00182
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шарма, А.Д.Е.В., Гилл, П.К., и Сингх, П. (2002). Выделение ДНК из сухих и свежих образцов растений, богатых полисахаридами. Завод Мол. биол. 20, 415а–415а.
Академия Google
Тамару Ю., Такани Ю., Йошида Т. и Сакамото Т. (2005). Решающая роль внеклеточных полисахаридов в устойчивости к высыханию и замораживанию у наземных цианобактерий Nostoc commune . Заяв. Окружающая среда. микробиол. 71, 7327–7333. doi: 10.1128/AEM.71.11.7327
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тиллетт, Д., и Нейлан, Б.А. (2000). Выделение ксантогенатной нуклеиновой кислоты из культивируемых и экологических цианобактерий. J. Phycol. 36, 251–258.
Академия Google
Уоррен-Роудс, К.А., Родс, К.Л., Пойнтинг, С.Б., Юинг, С.А., Лакап, Д.К., Гомес-Сильва, Б., и соавт. (2006). Гиполитические цианобактерии, сухой предел фотосинтеза и микробная экология в гипераридной пустыне Атакама. Микроб. Экол. 52, 389–398. doi: 10.1007/s00248-006-9055-9057
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wei, S.T.S., Lacap-Bugler, D.C., Lau, M.C.Y., Caruso, T., Rao, S., de los Rios, A., et al. (2016). Таксономическое и функциональное разнообразие почвенных и гиполитических микробных сообществ долины Майерс, Сухие долины Мак-Мердо, Антарктида. Перед. микробиол. 7:1642. doi: 10.3389/fmicb.2016.01642
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вежхос, Дж., DiRuggiero, J., Vitek, P., Artieda, O., Souza-Egipsy, V., Skaloud, P., et al. (2015). Стратегии адаптации эндолитических хлорофототрофов к выживанию в гипераридной и экстремальной солнечной радиационной среде пустыни Атакама. Перед. микробиол. 6:934. doi: 10.3389/fmicb.2015.00934
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вонг, Ф.К.Ю., Лакап, Д.К., Лау, М.С.Ю., Эйчисон, Дж.К., Коуэн, Д.А., и Пойнтинг, С.Б. (2010). Гиполитическое микробное сообщество кварцевого покрытия в высокогорных тундрах Центрального Тибета. Микроб. Экол. 60, 730–739. doi: 10.1007/s00248-010-9653-9652
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ян, Ю., Цинь, С., Чжао, Ф., Чи, X., и Чжан, X. (2007). Сравнение генов оболочки у одноклеточных и нитчатых цианобактерий. Комп. Функц. Геномика 2007:25751. дои: 10.1155/2007/25751
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Йошида, Т., и Сакамото, Т. (2009). Индуцированное водным стрессом накопление трегалозы и контроль трегалазы у цианобактерий Nostoc punctiforme IAM M-15. J. Gen. Appl. микробиол. 55, 135–145. doi: 10.2323/jgam.55.135
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Йошимура, Х.Ю., Камото, С.О., Сумурая, Ю.Т., и Хмори, М.О. (2007). Ген сигма-фактора группы 3, sigJ, ключевой регулятор устойчивости к высыханию, регулирует синтез внеклеточного полисахарида у цианобактерий Anabaena sp. штамм PCC 7120. DNA Res. 14, 13–24. doi: 10.1093/dnares/dsm003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжу, К., Косой, М., и Диттмар, К. (2014). HGTector: автоматизированный метод, облегчающий полногеномное обнаружение предполагаемых горизонтальных переносов генов. BMC Genomics 15:717. дои: 10.1186/1471-2164-15-717
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Утечка обновленных характеристик Roland SP-404 MkII — Tech
Пользователь Reddit опубликовал характеристики обновленной кассеты сэмплера Roland SP-404 MkII, раскрыв ее дополнительные функции и спекулятивную цену.
Культовый сэмплер рабочей станции включает в себя длинный список новых функций, если слухи верны — с 17 чувствительными к силе нажатия пэдами, 32-голосной полифонией, OLED-экраном для редактирования волновой формы и обновленной станцией эффектов.
Один пользователь на форуме Elektronauts перечислил все обновления после того, как обнаружил продукт, указанный на веб-сайте музыкального оборудования Сэма Эша, который был быстро удален. Оригинальному плакату удалось сделать быстрый скриншот просочившегося продукта.
Прочтите это далее: Roland TR-727 и 707 были добавлены в облако Roland
«В SP-404MKII есть все, что людям нравится в его предшественнике — с некоторыми серьезными обновлениями», — повторил пользователь из Sam Сайт Эш.
«Людям, которые создают музыку с помощью семплеров SP, нравится быстрый рабочий процесс, мобильность и тактильный подход, которые они предлагают», — говорится в просочившемся посте. «SP-404MKII еще больше расширяет этот опыт, сохраняя легкую, но прочную конструкцию, добавляя функции, которые упрощают создание в любом сценарии».
Дополнительные функции включают улучшенную скорость загрузки как для загрузки проекта, так и для запуска самой кассеты, в то время как обновление также позволяет использовать 16 ГБ памяти.
Кассета, наиболее известная благодаря использованию Lo-Fi-битмейкерами, также включает эффекты «Lo-Fi, Cassette Simulator и Resonator» в обновленном издании.
Читайте дальше: 10 самых влиятельных синтезаторов всех времен
«Обрабатывайте, переворачивайте и искажайте свои звуки с помощью легендарных эффектов SP и подборки новых классических произведений. Основные продукты SP, такие как Vinyl Simulator и DJFX Looper, теперь дополнены мощными дополнениями», — продолжает описание.
Предполагаемая цена нового Roland SP-404 MkII первоначально составляла 600 долларов, но с тех пор была обновлена до 499 долларов.
Джемма Росс — цифровой стажер Mixmag, следите за ней в Твиттере
Особенности — NASSENGER SP-1 | КОНИКА МИНОЛТА
Однопроходная технология обеспечивает сверхскоростную печать
Особенности
Оснащен специальными технологиями и функциями для минимизации времени простоя
Оснащен функцией компенсации ошибок, обнаруженных датчиками сопла печатающей головки.
Теперь можно равномерно регулировать плотность для каждого цвета на всех модулях печатающих головок благодаря функциям коррекции плотности с использованием датчиков изображения.
Недавно разработанный механизм автоматической очистки поверхности сопла модуля печатающей головки устраняет дополнительные трудозатраты и навыки, которые ранее требовались для очистки сопла, а также обеспечивает долгосрочную и стабильную печать.
Использует однопроходную технологию
NASSENGER SP-1 использует однопроходную технологию печати на ткани (носителе), подаваемой на ленту под фиксированные печатные секции.Результатом стал струйный принтер, качество печати которого сравнимо с трафаретными принтерами.
Недавно разработанный модуль печатающей головки
NASSENGER SP-1 оснащен новым модулем печатающей головки для однопроходной печати. Кроме того, наша запатентованная технология управления впрыском чернил вышла на новый уровень, который позволяет гибко регулировать чернила для малых, средних или больших размеров капель. Сверхвысокая скорость, высокая детализация и отличные результаты градации.
Удобное обслуживание
«Подъемная заслонка» обеспечивает прямой доступ к модулю печати в дополнение к «прогулке» для каждого цвета. Простое обслуживание означает повышенную производительность. Если модуль печатающей головки нуждается в замене, этот процесс прост и не требует специальных навыков. Точки попадания чернил можно легко отрегулировать с помощью датчиков изображения.
Сенсорная панель и дистанционное управление
Использует сенсорную панель для интуитивно понятного управления.Дистанционно вносите изменения в настройки производства и технического обслуживания с помощью планшета.
Чернила Konica Minolta для красивого проявления цвета
В дополнение к существующим чернилам на реактивных красителях были разработаны новые чернила на дисперсных красителях для однопроходной печати.